Что показывают карты генетического сцепления
Хромосомная теория
Хромосомная теория наследственности
Концепция данной теории заключается в том, что передача наследственной информации в ряду поколений осуществляется путем передачи хромосом, в которых в определенной линейной последовательности расположены гены.
Данная теория была сформулирована в начале XX века. Значительный вклад в ее развитие внес американский генетик Томас Морган.
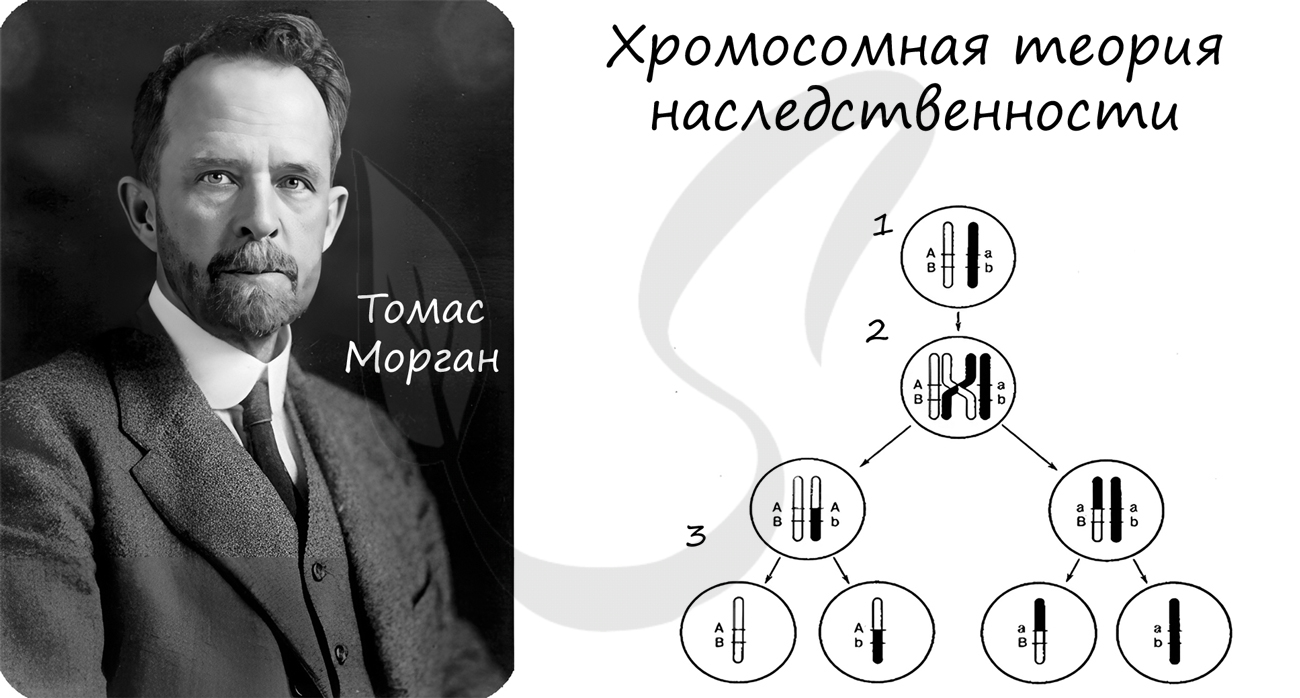
Рекомендую осознать и запомнить следующие положения хромосомной теории:
- Гены расположены в хромосомах в линейном порядке
- Каждый ген занимает в хромосоме определенное место — локус
- Гены, расположенные в одной хромосоме, образуют группу сцепления
- Сцепление генов может нарушаться в результате кроссинговера
- Частота кроссинговера между генами прямо пропорциональна расстоянию между ними
- Расстояние между генами измеряется в морганидах (1 морганида — 1% кроссинговера)
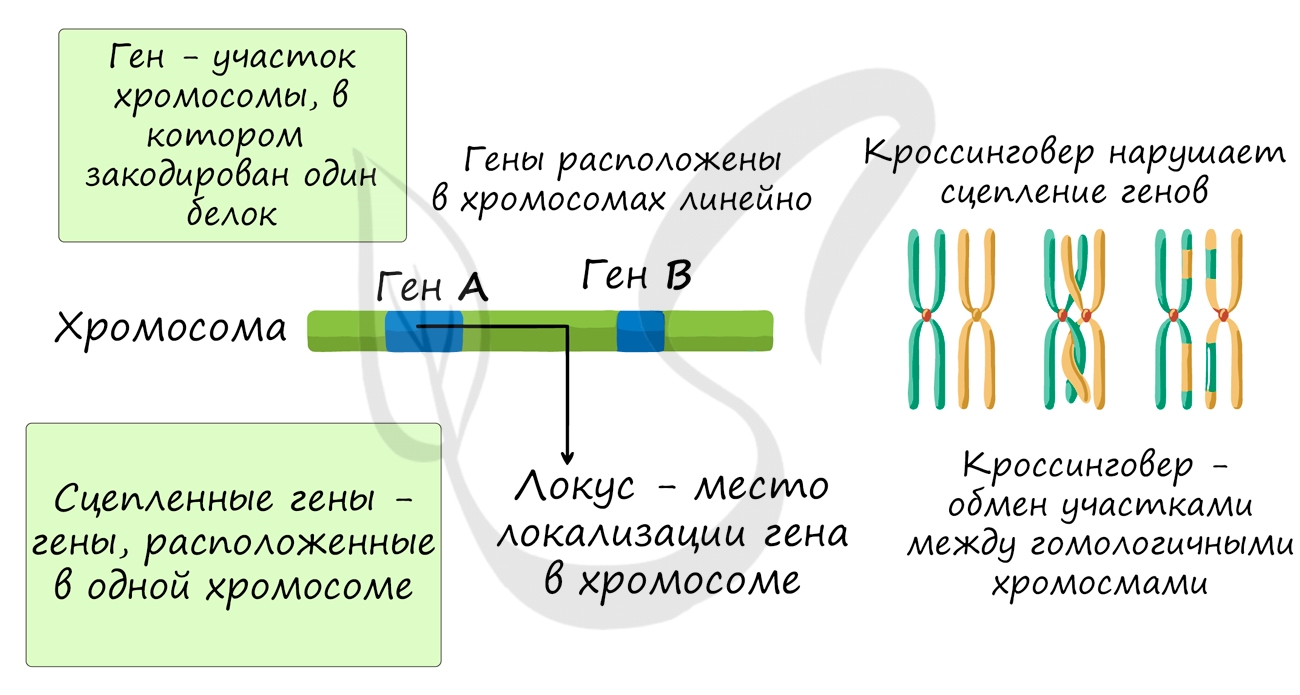
Группы сцепления
В предыдущей статье были раскрыты суть и применение в задачах III закона Менделя, закона независимого наследования, в основе которого лежат гены, расположенные в разных хромосомах. Но что если гены лежат в одной хромосоме? Такие гены образуют группу сцепления, в этом случае говорят о сцепленном наследовании.
Группа сцепления — совокупность всех генов, расположенных в одной хромосоме, вследствие чего они наследуются совместно. Число групп сцепления равно гаплоидному набору хромосом: у женщины 23 группы сцепления (23 пара — половые хромосомы XX), а у мужчины — 24 группы сцепления (X и Y представляют собой две отдельные группы).
Сцепление генов
Томас Морган в своих экспериментах изучал наследование признаков плодовых мушек дрозофил: серый (A) — черный (a) цвет тела, длинные (B) — зачаточные (b) крылья. В первом эксперименте Морган скрестил чистые линии плодовых мушек: серых с длинными крыльями (AABB) и черных с зачаточными (aabb).
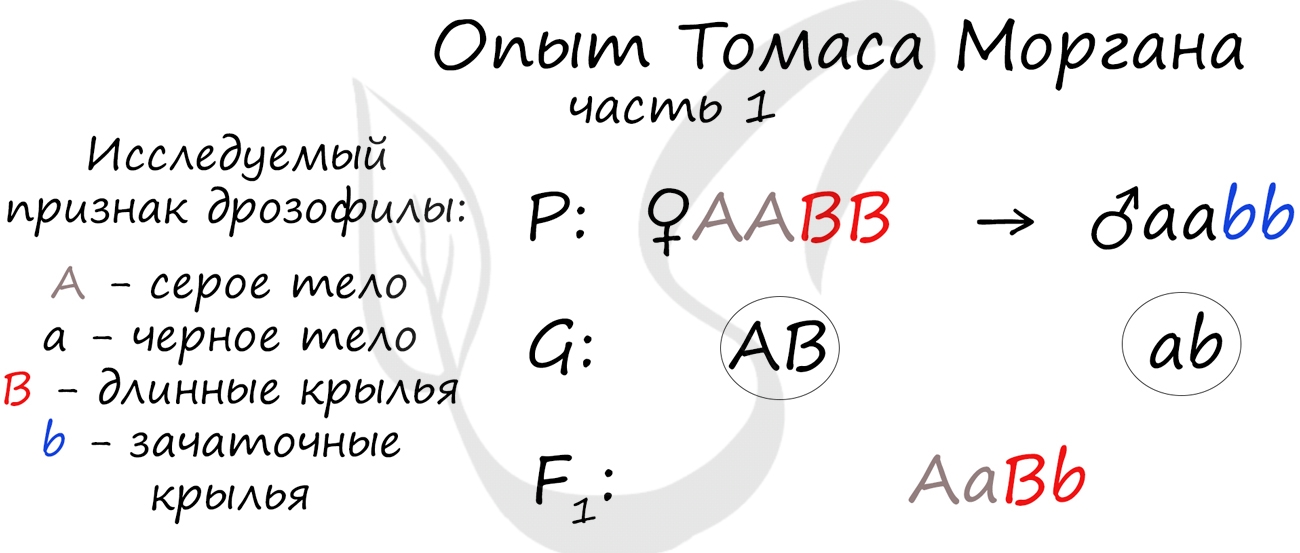
Только что вы видели первый закон Менделя (единообразия) в действии, правда, в несколько ином варианте — при дигибридном скрещивании. Но суть та же: в первом поколении все особи получаются единообразны по исследуемому признаку, с генотипом AaBb — с серым телом и длинными крыльями.
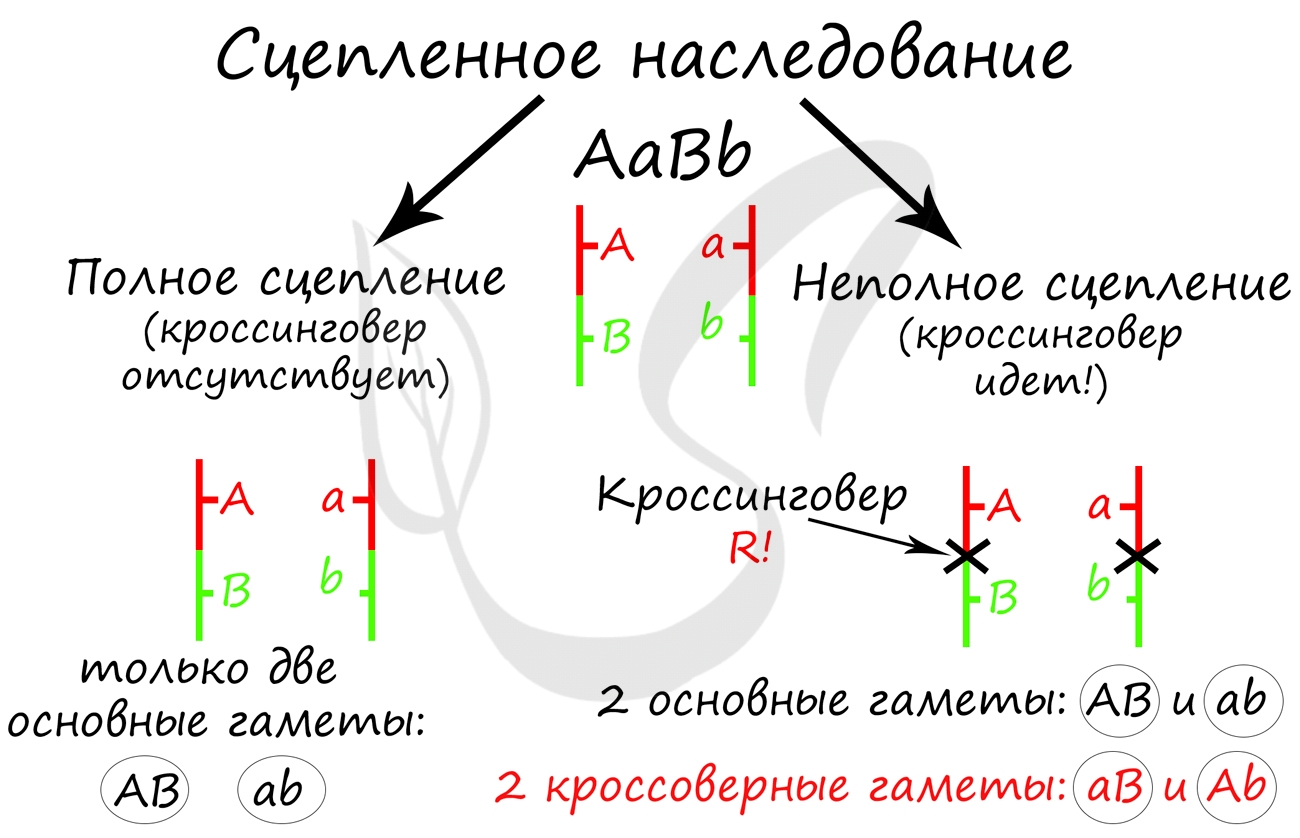
Далее Морган применил анализирующее скрещивание. Полученную в первом поколении дигетерозиготу (AaBb) он скрестил с черной особью с зачаточными крыльями (aabb). Результат весьма удивил Моргана и его коллег: помимо потомства с ожидаемыми фенотипами (серое тело + длинные крылья, черное тело + зачаточные крылья) были получены особи со смешанными признаками.

Потомство со смешанными признаками подразумевает под собой особи Aabb (серое тело + зачаточные крылья) и aaBb (черные тело + длинные крылья). Но откуда они могли взяться, если гены A и B находятся в одной хромосоме? Значит, образовались еще какие-то дополнительные гаметы, помимо AB и ab?
Объясняя полученные в потомстве фенотипы, которые содержали смешанные признаки, Томас Морган пришел к выводу, что между гомологичными хромосомами произошел кроссинговер, в результате которого образовались гаметы Ab, aB — кроссоверные гаметы.
Очевидно, что в данном случае расстояние между генами A и B было 17 морганид, так как каждой кроссоверной гаметы (соответственно и особей) образовалось по 8.5%. Не забывайте, что процент кроссинговера равен расстоянию между генами. Поскольку расстояние было 17 морганид = 17%, то на каждую из кроссоверных гамет приходится половина — 8.5%
Пример решения генетической задачи №1
«Катаракта и полидактилия у человека обусловлены доминантными аутосомными генами, расположенными в одной хромосоме. Гены полностью сцеплены. Какова вероятность родить здорового ребенка в семье, где муж нормален, жена гетерозиготна по обоим признакам, мать жены также страдала обеими аномалиями, а отец был нормален».
Очень важно обратить внимание на то, что «гены полностью сцеплены» — это говорит об отсутствии кроссинговера, и то, что мы заметили это, обеспечивает верное решение задачи.

Самое главное, что вам следует усвоить: поскольку гены полностью сцеплены (кроссинговер отсутствует), женщина с генотипом AaBb может образовать только два типа гамет — AB, ab. Кроссоверные гаметы (Ab, aB) не образуются. Всего возможных генотипов потомков получается два, из которых здоров только один — aabb. Шанс родить здорового ребенка в такой семье ½ (50%).
Пример решения генетической задачи №2
«Гены доминантных признаков катаракты и эллиптоцитоза локализованы в 1-й аутосоме. Гены неполностью сцеплены. Женщина, болеющая катарактой и эллиптоцитозом, отец которой был здоров, выходит замуж за здорового мужчину. Определите возможные фенотипы потомства и вероятность рождения больного обеими аномалиями ребенка в этой семье».
Ключевые слова в тексте этой задачи, на которые следует обратить внимание: «гены неполностью сцеплены». Это означает, что между ними происходит кроссинговер.
Генотип женщины остается неясен из текста задачи. Раз она больна, то он может быть: AaBb, AABB, AABb, AaBB. Однако в тексте дано то, что развеет сомнения: «отец которой был здоров». Если ее отец был здоров, то его генотип был aabb, значит он передал дочери гамету ab. Теперь становится очевидно, что генотип дочери AaBb — она дигетерозиготна.

В данном случае между генами A и B произошел кроссинговер, их сцепление нарушилось. В результате образовались кроссоверные гаметы Ab, aB — которые привели к образованию особей с со смешанными признаками (Aabb, aaBb). Вероятность рождения в этой семье ребенка, больного обеими аномалиями, составляет ¼ (25%).
Наследование, сцепленное с полом
Половые хромосомы X и Y определяют пол человека. Генотип XX характерен для женщин, а XY — для мужчин. Мужская Y-хромосома не содержит аллелей многих генов, которые есть в X-хромосоме, вследствие этого наследственными заболеваниями, сцепленными с полом, чаще болеют мужчины.
Природа, несомненно, бережет женских особей. Женщины имеют две гомологичные хромосомы XX, и если ген наследственного заболевания попал в одну из X-хромосом, то чаще всего в другой X-хромосоме окажется «здоровый» ген, доминантный, которой подавит действие рецессивного гена. С генетической точки зрения, женщина будет носительницей заболевания, может его передать по поколению, но сама болеть не будет.
У мужчин если ген заболевания оказался в X-хромосоме, то не проявиться он не может. Именно по этой причине мужчины чаще страдают дальтонизмом, гемофилией и т.д.
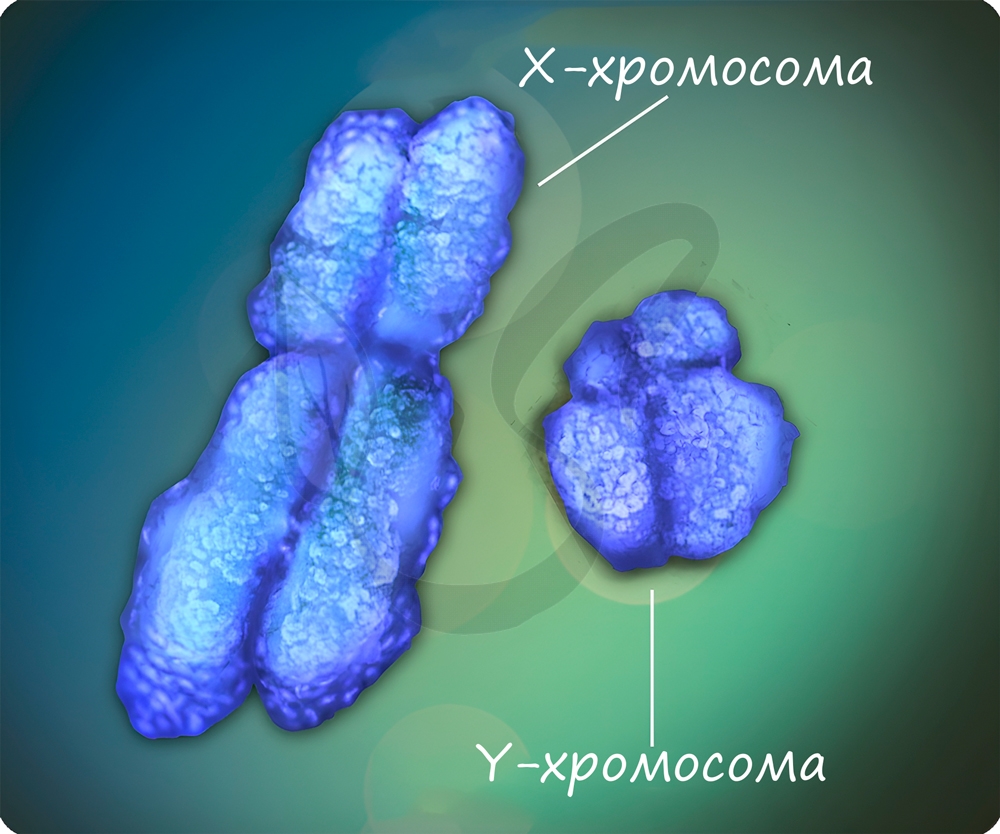
Не у всех организмов особь мужского пола характеризуется набором хромосом XY, а женского — XX. У пресмыкающихся, птиц, бабочек женские особи имеют гетерогаметный пол- XY, а мужские — XX. То же самое относится к домашним курам: петух — XX, курица — XY.
Решим несколько задач по теме наследования, сцепленного с полом. Речь в них будет идти о сцепленных с полом признаками — признаками, гены которых лежат не в аутосомах, а в гетеросомах (половых хромосомах).
Пример решения генетической задачи №3
«Рецессивный ген дальтонизма располагается в X-хромосоме. Женщина с нормальным зрением (отец был дальтоник) выходит замуж за мужчину с нормальным зрением, отец которого был дальтоником. Определите возможные фенотипы потомства».
Подробности о родословной важны и помогают заполнить белые пятна. Если отец женщины был дальтоником (X d Y), то очевидно, что он передал ей хромосому X d , так как от отца дочери всегда передается X-хромосома. Значит женщина гетерозиготна по данному признаку, а у мужчины возможен лишь один вариант здорового генотипа — X D Y. То, что его отец был дальтоником несущественно, ведь отец всегда передает сыну Y-хромосому.

Возможные фенотипы потомства:
- X D X D , X D X d — фенотипически здоровые девочки
- X D Y — здоровый мальчик
- X d Y — мальчик, который болен дальтонизмом
Пример решения генетической задачи №4
«Гипоплазия зубной эмали наследуется как сцепленный с X-хромосомой доминантный признак, шестипалость — как аутосомно-доминантный. В семье, где мать шестипалая, а у отца гипоплазия, родился пятипалый здоровый мальчик. Напишите генотипы всех членов семьи по данным признакам. Возможно ли у них рождение ребенка с двумя аномалиями одновременно?»
Ответ на вопрос: «Каковы генотипы матери и отца?» — лежат в потомстве. Пятипалый здоровый мальчик имеет генотип aaX b Y. Чтобы сформировался такой генотип, от матери должна прийти гамета aX b , а от отца — aY. Выходит, что единственно возможный генотип матери — AaX b X b , а генотип отца — aaX B Y.

Рождение ребенка с двумя аномалиями возможно — AaX B X b , вероятность такого события ¼ (25%).
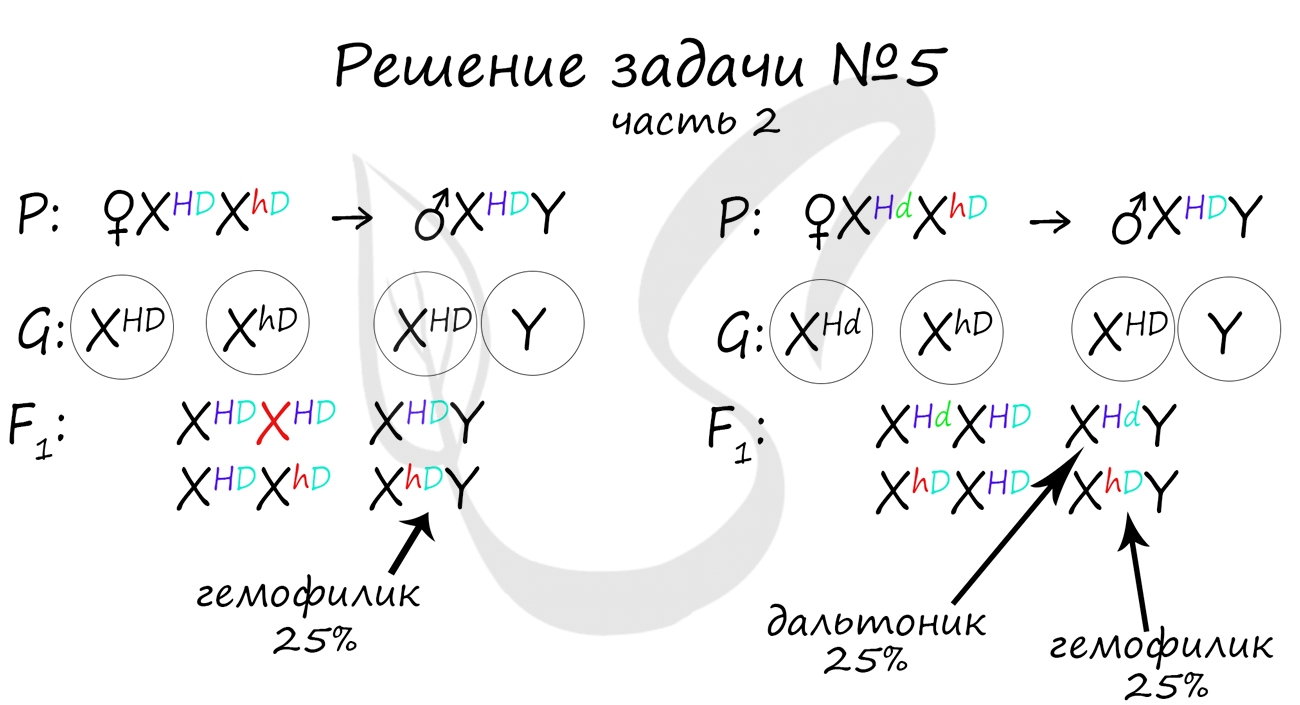
Пример решения генетической задачи №5
«Рецессивные гены, кодирующие признаки дальтонизма и гемофилии, сцеплены с X-хромосомой. Мужчина с нормальным цветовым зрением и гемофилией женится на здоровой женщине, отец которой был дальтоником, но не гемофиликом. Известно, что мать женщины была гомозиготна по исследуемым признакам. Какое потомство получится от брака их дочери со здоровым мужчиной?»
Генотип мужчины вопросов не вызывает, так как единственный возможный вариант — X hD Y. Генотип женщины дает возможность узнать ее отец (X Hd Y), который передал ей гамету X Hd (отец всегда передает дочке X хромосому, а сыну — Y), следовательно, ее генотип — X HD X Hd
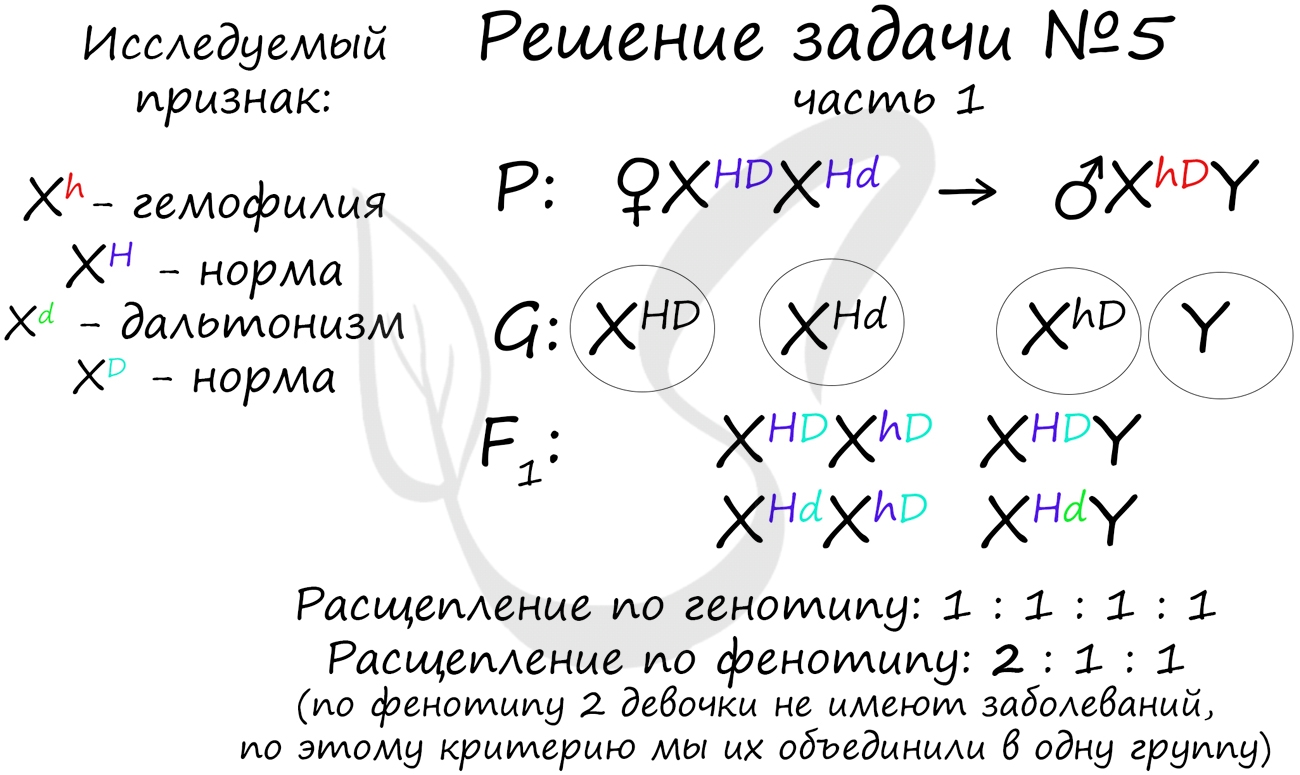
Как оказалось, возможны два варианта генотипа дочери: X HD X hD , X Hd X hD . Генотип здорового мужчины X HD Y. Следуя логике задачи, мы рассмотрим два возможных варианта брака.
Не забывайте, что на экзамене схема задачи не является ответом. Ответ начинается только после того, как вы напишите слово «Ответ: . «. В ответе должны быть указаны все фенотипы потомства, их описание, что возможно покажется рутинными при большом числе потомков, но весьма приятным, если вы верно решили задачу и получили за нее заслуженные баллы 
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Генетические карты сцепления: общие сведения
Генетическая карта сцепления (карта сцепления, генетическая карта, linkage map, genetic map) [лат. charta — бумага, грамота] — одномерная схема взаиморасположения локусов генов (генетических маркеров) в группах сцепления (на индивидуальных хромосомах) данного организма с указанием расстояний между ними в парах нуклеотидов, определенных с помощью секвенирования, или в сантиморганах, установленных по частоте кроссинговера.
Генетические карты сцепления правильно отражают порядок расположения генетических маркеров на хромосомах, однако полученные при этом значения расстояний между ними не соответствуют реальным физическим расстояниям. Обычно данный факт связывают с тем, что эффективность рекомбинации между хроматидами на отдельных участках хромосом может сильно различаться. В частности, она подавлена в гетерохроматиновых участках хромосом. С другой стороны, в хромосомах часто встречаются «горячие точки» рекомбинации. Использование частот рекомбинации для построения физических генетических карт без учета этих факторов будет приводить к искажениям реальных расстояний между генетическими маркерами.
Таким образом, генетические карты сцепления являются наименее точными из всех имеющихся типов генетических карт, и их можно рассматривать только в качестве первого приближения к реальным физическим картам. Тем не менее, на практике именно они позволяют локализовать сложные генетические маркеры (например ассоциированные с симптомами заболевания) на первых этапах исследования и дают возможность их дальнейшего изучения. Необходимо помнить, что в отсутствие кроссинговера все гены, находящиеся на индивидуальной хромосоме, передавались бы от родителей потомству вместе, поскольку они физически сцеплены друг с другом. Поэтому индивидуальные хромосомы образуют группы сцепления генов, и одной из первых задач построения генетических карт сцепления является отнесение исследуемого гена или последовательности нуклеотидов к конкретной группе сцепления. В табл. II.4 перечислены современные методы, которые, по данным В.А. МакКьюзика, наиболее часто использовались для построения генетических карт сцепления до конца 1990 г.
Впервые ген был отнесен к определенной хромосоме в 1911 г., когда E.B. Wilson на основании анализа родословной смог отнести ген дальтонизма к X-хромосоме.
В последующем, по мере открытия новых генов, встала задача определения не только их хромосомной локализации, но и порядка расположения на хромосоме и расстояния между ними. Решение этой задачи стало возможно после открытия явления генетической рекомбинации. Появилась возможность построения генетических карт, т.е. карт, показывающих порядок расположения генов и относительных расстояний между ними. Единицей измерения расстояний при генетическом картировании является один сантиморган (сМ), т.е. такое расстояние, при котором вероятность рекомбинации между генами равно 1%.
Наиболее существенным недостатком генетических карт является их неаддитивность, то есть, если известно, что расстояние между генами A и B равно 3 сМ, B и C — 5 сМ, то это вовсе не значит, что расстояние между A и C равно 8 сМ. Одной из причин этого является то, что в разных районах генома рекомбинация происходит с разной частотой.
С тех пор методы генетического картирования совершенствовались, но основной принцип оставался неизменным — гены картировались на основании анализа распределения между родственниками. В определенном смысле венцом такого подхода стала карта человеческого генома, опубликованная в 1987 г. H. Donis-Keller с соавт. [ Donis_Koler H. e. a., 1987 ]. Получение генетических карт было значительно ускорено технологией использования маркеров на основе коротких динуклеотидных повторов, что позволило строить генетические карты с недостижимым прежде разрешением. Например, карта сцепления на основе высокополиморфных динуклеотидных маркеров, полученная во многом усилиями Genethon, в настоящее время состоит из 2066 маркеров, расположенных в среднем на расстоянии 2,9 сМ друг от друга [ Gyapay G. e. a., 1994 ].
Такая информация недостаточна для решения тех задач, которые стоят перед молекулярной генетикой в настоящее время. Для изучения молекулярных основ наследственности чрезвычайно важно знать абсолютные расстояния между генетическими элементами. Таким образом, встает задача построения физической карты генома или его участков, то есть такой карты, которая давала бы абсолютные расстояния между генетическими маркерами.
Карта генетического сцепления генома человека
2. Карта генетического сцепления генома человека
Генетика человека существенно продвинется вперед с появлением полной карты сцепления. Такая карта будет состоять из маркеров ПДРФ, равномерно разбросанных по всем хромосомам, покрывая в целом около 3300 сантиморганид.
За последние несколько лет в этом направлении был сделан значительный шаг вперед. В конце 1987 г. Донис-Келлер сообщил о построении карты для 403 локусов генома человека, распределенных по всем группам сцепления соответственно числу хромосом. В это же время Уайт распространил буклеты, содержащие 255-локусную карту, с группами сцепления, охватывающими 17 из 23 хромосом. Так как перекрывание между наборами маркеров, изученными в этих двух группах, невелико, возможно построение 600-локусной суммарной карты. Учитывая, что в настоящее время этими и другими группами исследователей картируется множество дополнительных ПДРФ-маркеров, представляется вполне реальным, что вскоре в нашем распоряжении окажется карта для более чем 1000 ПДРФ. Эта карта будет иметь в среднем по одному маркеру на каждые 3 см.
Такая плотная карта ПДРФ-маркеров чрезвычайно удобна для анализа человеческого генома.
1. Все без исключения семьи оказываются полностью информативными для анализа сцепления. Когда в анализе сцепления используются случайные и некартированные ПДРФ-маркеры, мейозы, происходящие у индивидов, гомозиготных по одному из них, не являются информативными для анализа сцепления ПДРФ-маркера с интересующим нас заболеванием. Наличие плотной карты позволяет, однако, выбирать в любом районе несколько близко расположенных ПДРФ таким образом, чтобы все индивиды были заведомо гетерозиготными хотя бы по одному из них. Получение такой информации позволяет осуществить анализ полилокусного сцепления с помощью компьютера.
2. Наследование локуса, ответственного за заболевание, можно проследить более строго, используя фланкирующие маркеры. Бели между маркерами, расположенными по обе стороны от предполагаемого локуса, ответственного за болезнь, рекомбинация не наблюдается, можно быть уверенным, что данный локус, расположенный между ними, наследовался совместно с этими маркерами. В действительности, в генетике давно уже считается, что анализ по таким тройным перекрестам много точнее, чем по двойным. Повышенная точность упрощает процедуру принятия или отклонения гипотезы, и таким образом, уменьшает необходимое для анализа количество семей.
3. Можно одновременно проследить наследование нескольких разных районов генома. Это позволяет установить сцепление в случае генетической гетерогенности заболевания.
Нам представляется, что в будущем анализ сцепления первоначально будет основан на использовании стандартного набора приблизительно 150–200 высокоинформативных ПДРФ-маркеров, распределенных по всему генному, с интервалом около 15–20 см. После того как все изучаемые родословные окажутся генотипировэнными по каждому маркерному локусу, будет проведен компьютерный анализ, тестирующий каждый такой участок на наличие шансов за сцепление. Любой фрагмент, предположительно сцепленный с изучаемым признаком, будет изучен затем более подробно с использованием плотной карты ПДРФ, например с плотностью один маркер на каждые 3 см. Это позволит извлечь всю информацию, заключенную в родословной, и увеличит величину лод-балла. Естественно, при планировании такого анализа вполне уместен вопрос: сколько семей необходимо, чтобы при наличии редкой карты ПДРФ уловить хотя бы намек на сцепление, а для плотной карты ПДРФ иметь возможность доказать с определенностью наличие сцепления?
3. Планирование анализа сцепления для болезней с простым типом наследования
Под простым типом наследования мы понимаем, что болезнь: 1) наследуется как классический менделеевский доминантный или рецессивный признак, проявляющий полную пенетрантность, и 2) вызвана мутациями в одном локусе. Данные точных эпидемиологических исследований заболевания обычно дают основания судить о том, справедливо первое условие или нет. В противоположность этому второе условие обычно следует принимать на веру: не существует такого мощного метода для доказательства того, что мутации по одному локусу являются причиной данного заболевания, и в то же время существует много поводов усомниться в этом. Но к данному вопросу мы вернемся позже, при обсуждении генетической гетерогенности.
Болезнь Гентингтона и муковисцидоз являются яркими примерами признаков с доминантным и рецессивным типом наследования, соответственно. Оказывается, что в обоих случаях болезнь – следствие мутаций единичного локуса.
Для заболевания с простым типом наследования относительно легко построить карту сцепления. Рассчитав сумму ELOD-баллов, можно прямо оценить требуемую величину выборки. На рис. А, Б показано число ядерных семей с двумя или тремя пораженными, которое требуется, чтобы гарантировать 50%-ную вероятность успеха при картировании доминантного или рецессивного признака. Это число является функцией плотности расположения используемых ПДРФ-маркеров. Рисунок демонстрирует количество семей, необходимое 1) в случае, когда несколько маркеров рестрикционного полиморфизма используются по отдельности, и 2) если задействован весь потенциал карты ПДРФ, т.е. используются фланкирующие маркеры. Как следует из рис. 1, необходимое количество семей вполне эффективно при наличии геномной карты ПДРФ, отстоящих друг от друга на расстоянии 20–30 см.
Далее мы рассмотрим признаки с более сложным типом наследования и обсудим проблемы, возникающие при их картировании с помощью анализа сцепления, и пути их преодоления.
Что показывают карты генетического сцепления
Анализ сцепления — метод картирования генов, использующий семейные исследования для определения связи между двумя генами при их передаче от одного поколения другому. Для того чтобы решить, что два локуса сцеплены и, если да, то как сильно, мы основываемся на двух видах информации.
Во-первых, мы устанавливаем, значимо ли отклоняется частота рекомбинации Q между двумя локусами от 0,5; определение сцепления между двумя локусами эквивалентно выяснению того, отличается ли доля рекомбинаций между ними от 0,5 ожидаемой для несцепленных локусов.
Во-вторых, если доля рекомбинаций менее чем 0,5, нужно оценить ее наилучшим образом, поскольку это покажет, насколько тесно сцеплены локусы. В обоих случаях применяют статистический метод отношения правдоподобия. Правдоподобие — меры вероятности, шансы — отношения правдоподобия. Отношения правдоподобия рассчитывают следующим образом.
Изучают фактические семейные данные, подсчитывают число детей, имеющих рекомбинацию между локусами и, наконец, вычисляют правдоподобие (вероятность) наблюдаемого значения Q в интервале от 0 до 0,5. Затем вычисляют вторую вероятность, основанную на гипотезе, что два локуса не сцеплены, т.е. Q=0,50. Мы рассматриваем отношение правдоподобия наблюдаемых в семейных данных значений Q и правдоподобия при условии отсутствия сцепления между локусами и тем самым получаем отношение шансов:
1) Правдоподобие данных, если локусы сцеплены с некоторым коэффициентом Q
2) Правдоподобие данных, если локусы не сцеплены (Q = 0)
Вычисленные отношения шансов для значений Q обычно представляют в форме десятичных логарифмов и называют LOD-оценкой (Z) логарифма шансов (log of the odds). (Использование логарифмов позволяет объединять данные, полученные из разных семей простым сложением).
Анализ сцепления менделирующих болезней, основанный на моделях (прототипах)
• Анализ сцепления называют модельным (или параметрическим), если допускают, что существует конкретный тип наследования (аутосомно-доминантный, аутосомно-рецессивный или Х-сцепленный), объясняющий наследование признака.
• Анализ LOD-оценки допускает картирование генов, мутации в которых вызывают болезни, передающиеся по менделирующему типу.
• LOD-оценка дает:
— наилучшую оценку частоты рекомбинации (Qmax) между маркерным локусом и локусом болезни;
— оценку того, насколько хорошо подтверждается сцепление этой величиной Qmax. Величины LOD-оценки более 3 считают надежным подтверждением.
• Сцепление с конкретным значением Qmax локуса гена болезни и маркера с известной физической позицией подразумевает, что локус гена болезни должен располагаться около маркера.
Отношение шансов важно в двух смыслах. Во-первых, оно обеспечивает статистически правильный метод для использования семейных данных при оценке частоты рекомбинации между локусами. Дело в том, что статистическая теория гласит, что величина, дающая наибольшую величину Z, фактически, наилучшая оценка доли рекомбинации, которую можно сделать, основываясь на имеющихся данных. Эта величина называется Qmax. Если Q отличается от 0,50, мы получаем подтверждение сцепления.
Тем не менее хотя Qmax — наилучшая оценка Q, насколько она хороша? Отношение шансов отвечает и на этот вопрос, поскольку, гем выше велигина Z, тем лугше оценка Qmax. Положительные величины Z (шансы >1) при данной Q указывают, чтобы два локуса сцеплены, тогда как отрицательные (шансы 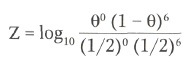
Максимальная величина Z равна 1,81, встречающаяся, когда Q=0, и предполагает, но не уверенно, наличие сцепления, поскольку значение Z — положительное, но меньше 3.
Объединение информации LOD-оценки из разных семей
Таким же образом, каждый мейоз в семье, производящей нерекомбинантное или рекомбинантное потомство — независимый случай, так же независимы и мейозы, происходящие в других семьях. Следовательно, мы можем перемножить вероятности в числителе и знаменателе отношений правдоподобия отдельных семей. Аналогичное, но более удобное вычисление — сложить десятичные логарифмы (log10) всех вычисленных отношений правдоподобия, формируя общую Z-оценку для всех семей.
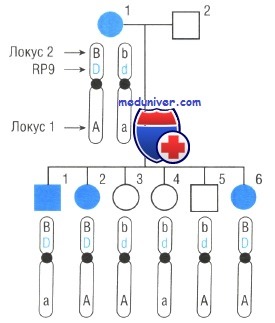 Родословная наследования пигментного ретинита
Родословная наследования пигментного ретинита
В случае пигментного ретинита на рисунке, предположим, что изучались две других семьи и в одной не обнаружена рекомбинация между локусом 2 и пигментным ретинитом у четырех детей, а в третьей нет рекомбинации у пяти детей. Для каждой семьи рассчитаны индивидуальные LOD-оценки, затем сложенные вместе. В этом случае, можно было бы говорить, что ген пигментного ретинита в этой группе семей сцеплен с локусом 2.
Поскольку хромосомная позиция полиморфного локуса 2 известна — 7р14, пигментный ретинит в этой семье может картироваться в области вокруг 7р14, рядом с локусом RP9, уже определенным для одной из форм аутосомно-доминантного пигментного ретинита.
Если некоторые анализируемые семьи имеют пигментный ретинит из-за мутаций в другом локу-се, оценки LOD между семьями будут расходиться: некоторые будут показывать небольшую положительную, другие — резко отрицательную оценку. Можно все еще складывать Z-оценки вместе, но результат даст быстрый спад в общей LOD-оценке. Таким образом, при анализе сцепления, включающем более одной семьи, не заподозренная гетерогенность локуса может скрывать сцепление, существующее в подмножестве семей.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Как ученые расшифровывают геномы и зачем это нужно? Отвечаем в 9 карточках
В последние годы ученые постоянно объявляют о расшифровке геномов тех или иных видов. ITMO.NEWS и ученый Международного научного центра SCAMT Алексей Комиссаров в карточках объясняют: что такое ДНК, как с ее помощью изучают историю животных и в чем отличие ДНК-теста от геномного исследования.

Иллюстрации: Дмитрий Лисовский, ITMO.NEWS
По пунктам:
- Начнем с простого: что такое ДНК?
- Говорят, ДНК можно получить из волоса, слюны, крови. А каким образом?
- А есть еще геном. Это одно и то же?
- Я все время слышу: расшифровали геном того, расшифровали геном сего. А кто его зашифровывает?
- Говорят, по геному можно проследить генетическую историю. Это что-то типа ДНК-теста?
- А как ученые берут кровь или шерсть у современного льва, а получают данные о его предках?
- Мы говорим, что взяли кость тигра раннего палеолита и по ней определили какие-то вещи про целую популяцию. Но как мы отделяем эту информацию от той, что была присуща именно вот этому самому тигру?
- А как мы проверяем наши выводы? Ведь не всегда мы можем выкопать пещерного предка современных животных и удостовериться, что все сходится.
- Зачем нам все это?
Начнем с простого: что такое ДНК?

Каждый более или менее знает, что есть белки, жиры и углеводы. Но еще у нас в каждой клетке есть ДНК, дезоксирибонуклеиновые кислоты, которые отвечают за хранение информации. Для геномного биоинформатика ДНК ― это прежде всего один из главных языков биологии, который состоит всего лишь из четырех букв: A, T, G и C. Эти буквы являются сокращениями имен четырех азотистых оснований, из которых состоит ДНК: аденин (А), цитозин (C), гуанин (G) и тимин (Т). ДНК можно сравнить с компьютерной программой, очень сложной, запутанной, со множеством ошибок и костылей, но, тем не менее, она работает.
Говорят, ДНК можно получить из волоса, слюны, крови. А каким образом?

ДНК содержится почти во всех клетках организма, исключение — эритроциты, которые в зрелом состоянии теряют ядро, чтобы было легче переносить кислород. Поэтому биологический материал для выделения ДНК может быть разнообразен. Процесс выделения ДНК состоит из четырех этапов: разрушение мембраны клеток для высвобождения ДНК; очистка от связанных с ДНК белков; очистка от разного рода примесей; растворение ДНК для хранения. ДНК можно выделить и в домашних условиях, но тогда ДНК будет не очень чистой и ее будет сложно использовать для каких-либо научных исследований. От качества этих процедур будет зависеть и полученная из нее информация. Поэтому необходима лабораторная точность работы специалистов, молекулярных биологов. Проще всего ДНК выделять из крови, потому что это легкий для получения биоматериал. Хотя в эритроцитах ДНК нет, в крови плавает огромное количество других клеток — белых кровяных телец, так что и ДНК из них получается много. Из слюны или кусочков эпителия во рту ― уже меньше, из волос ― еще меньше. Например, чтобы прочитать весь геном достаточно точно, необходима кровь, а не слюна. А для того, чтобы сделать какой-нибудь ДНК-тест, где точность не очень важна ― достаточно и слюны.
А есть еще геном. Это одно и то же?

ДНК — это название молекулы, которая хранит наследственную информацию. Геном ― это совокупность всей ДНК организма со всеми записанными в ней особенностями конкретного вида или даже индивида. Поэтому можно говорить о геноме человека вообще, а можно — о геноме конкретных Васи или Кати. На физическом уровне геном разделен на хромосомы, в случае человека — 23 пары хромосом, 23 от мамы, и 23 от папы, всего 46. Когда организм начинает расти после оплодотворения, в каждой клетке копируется этот набор, но иногда это происходит с небольшими ошибками. Это называется соматическими мутациями. Иногда эти ошибки могут быть весьма критическими и приводить к разным заболеваниям.
Я все время слышу: расшифровали геном того, расшифровали геном сего. А кто его зашифровывает?

Его никто не зашифровывал, но это слово хорошо передает ощущения от работы с геномными данными. Если продолжать аналогию с геномом как с очень сложной программой, можно сказать, что она не только очень сложная, но и очень плохо написана. И кроме собственно четырех букв A, C, G и Т, он содержит много дополнительных уровней кодирования информации, которые не обязательно будут наследоваться и могут меняться в процессе жизни организма. Это часто называют эпигеномом, который изучает эпигенетика. Вся эта неимоверная сложность и создает ощущение расшифровки. Помимо этого, злую шутку здесь сыграл не очень корректный перевод с английского, где использовали слово decoding и encoding, декодировали и закодировали. Код — это просто система условных обозначений, не предполагающая никакого секрета, никакой защиты от взлома. Любой человеческий язык — это код, система дорожных знаков — это код. Шифр — это код, намеренно защищенный от взлома. Но, конечно, в английских терминах меньше романтики, чем в слове расшифровали.
Говорят, по геному можно проследить генетическую историю. Это что-то типа ДНК-теста?

Под ДНК-тестом часто имеют в виду анализ только некоторых небольших участков генома, вариации в которых имеют какой-то известный эффект. В геномных исследованиях ученые работают с гораздо большим количеством ДНК, в идеале со всей доступной информацией. Это называется полногеномными исследованиями. Но даже генетических тестов, направленных на выяснения значения только некоторых фрагментов генома, часто достаточно для того, чтобы проследить генетическую историю или оценить степень родства между двумя людьми. Это возможно, во-первых, благодаря тому, что у нас в геноме есть фрагменты, которые очень вариабельны и отличаются у разных людей, и, во-вторых, благодаря математике.
А как ученые берут кровь или шерсть у современного льва, а получают данные о его предках?

Есть очень сложные математические алгоритмы, которые позволяют по генетическим данным найти наиболее вероятный сценария развития событий: когда происходили мутации отдельных фрагментов, которые привели к образованию того генома, который мы видим сейчас. Своего рода, математическая машина времени. Ученые ИТМО недавно опубликовали программу, направленную как раз на решение проблемы — как наиболее точно заглянуть в прошлое генома. Одним из самых захватывающих расширений этого подхода является добавление еще и географических точек. Тогда мы можем не только смоделировать, как происходили изменения в геноме, ни и посмотреть, как отдельные популяции с этими геномными вариациями перемещались из одной точки в другую.
Мы говорим, что взяли кость тигра раннего палеолита и по ней определили какие-то вещи про целую популяцию. Но как мы отделяем эту информацию от той, что была присуща именно вот этому самому тигру?

Мы стараемся получить модель, которая наиболее правдоподобно описывает сценарий развития событий в прошлом. Чтобы сделать модель более точной, одного образца часто недостаточно, и чем больше образцов у нас есть, тем более точной становится наша модель. У каждого из нас очень много редких генетических вариантов, бывают и варианты, которые присущи только нам. И если у нас есть уже несколько образов, то такие индивидуальные варианты ученые отфильтровывают именно затем, чтобы они не мешали анализу. Так как единицей эволюции является популяция, а не отдельный индивид.
А как мы проверяем наши выводы? Ведь не всегда мы можем выкопать пещерного предка современных животных и удостовериться, что все сходится.

Хорошие модели обладают предсказательной способностью. Проверить проще всего новыми данными, которые не должны противоречить модели, но случается, что они противоречат, и тогда модель приходится пересчитывать. Сейчас мы наблюдаем очень красивую иллюстрацию мутаций, эволюции и вот этого всего на примере геномов коронавируса SARS-CoV-2. Настолько подробных данных об эволюции отдельного вида в реальном времени у человечества еще никогда не было. И появляется все больше данных и для остальных геномов. Со временем модели будут все лучше и лучше, а чем больше данных ― тем лучше модели.
Зачем нам все это?

Кроме того, что это захватывающе любопытно, это имеет множество практических применений во всех сферах деятельности человека. Если продолжать рассуждать об исследовании истории предков, то есть математический аппарат для поиска так называемых событий бутылочного горлышка, когда размер популяции по каким-то причинам резко сократился. Поиск таких событий, своего рода, геномная археология, может дать нам подсказки, как таких событий избежать. Это важно особенно сейчас, когда многие виды животных бесследно исчезают с лица земли.
Современные методы построения генетических карт сцепления
Число картированных локусов
Гибридизация соматических клеток
Гибридизация in situ
Семейный генетический анализ сцепления
Определение эффекта дозы
Использование хромосомных аберраций
Сегрегация генов, индуцированная облучением
Гибридизация соматических клеток. Одним из наиболее популярных методов отнесения генетического маркера (функционально активного гена) к конкретной группе сцепления является гибридизация (слияние друг с другом) соматических клеток разных биологических видов организмов, один из которых – исследуемый. У межвидовых гибридов соматических клеток в процессе культивирования происходит утрата хромосом преимущественно одного из биологических видов. Потеря хромосом носит, как правило, случайный характер, и образующиеся клоны клеток содержат оставшиеся хромосомы в разных сочетаниях. Анализ клонов, содержащих разные наборы хромосом исследуемого вида, позволяет определить, с какой из этих оставшихся хромосом ассоциирована экспрессия исследуемого маркера, и, следовательно, локализовать ген на конкретной хромосоме.
Гибридизация in situ. Метод гибридизации in situ также широко используется для картирования последовательностей нуклеотидов на хромосомах. С этой целью препараты фиксированных хромосом гибридизуют (инкубируют при повышенной температуре с последующим охлаждением) с исследуемыми последовательностями нуклеотидов, меченными радиоактивной, флуоресцентной или иной меткой. После отмывания несвязавшейся метки оставшиеся меченые молекулы нуклеиновых кислот оказываются ассоциированными с участками хромосом, содержащими последовательности, комплементарные исследуемым меченым последовательностям нуклеотидов. Полученные гибриды анализируют с помощью микроскопа либо непосредственно, либо после авторадиографии. Для этой группы методов характерна более высокая разрешающая способность, чем для гибридизации соматических клеток, поскольку они позволяют локализовать изучаемые последовательности нуклеотидов на хромосомах. По мере выполнения программы «Геном человека» в руках исследователей появляется все больше изолированных последовательностей нуклеотидов, которые можно использовать в качестве зондов для гибридизации in situ. В связи с этим данные методы по частоте использования в последнее время прочно выходят на первое место. Наиболее популярной оказывается группа методов, получивших название флуоресцентной гибридизации in situ (fluorescence in situ hybridization – FISH), при проведении которой используются полинуклеотидные зонды, содержащие флуоресцентную метку. В частности, в 1996 г. было опубликовано >600 работ, в которых описано использование этого метода.
Семейный генетический анализ сцепления. Эта группа методов часто используется в медицинской генетике для выявления связи (сцепления) между симптомами заболевания, вызываемого мутацией в неизвестном гене, и другими генетическими маркерами. В данном случае в качестве одного из генетических маркеров выступают сами симптомы заболевания. В геноме человека обнаружено большое количество полиморфизмов, в том числе ПДРФ (см. разделы 11.1 и 11.2). ПДРФ распределены более или менее равномерно в геноме человека на расстоянии 5–10 сМ друг от друга. Чем ближе индивидуальные полиморфные локусы расположены к гену, ответственному за заболевание, тем меньше вероятность их разделения при рекомбинации в мейозе и тем чаще они будут встречаться вместе у больного индивидуума и вместе передаваться от родителей потомству. Клонировав протяженный участок генома, включающий соответствующий полиморфный маркер (его отбор из клонотеки геномной ДНК проводят с помощью зонда), можно одновременно вместе с ним с большой вероятностью выделить ген, вызывающий наследственное заболевание. Такие подходы были, в частности, успешно применены для проведения семейного анализа и выделения соответствующих генов при мышечной дистрофии Дюшенна, кистозном фиброзе почек (муковисцидозе) и миотонической дистрофии. Информативность отдельных ПДРФ генома человека зависит от уровня их гетерозиготности в исследуемой популяции. Мерой информативности ПДРФ как генетического маркера по предложению Д. Ботштейна и соавторов (1980 г.) принято считать значение содержания полиморфной информации PIC (polymorphism information content), которое представляет собой отношение числа скрещиваний, в которых хотя бы у одного из родителей исследуемый полиморфный маркер находится в гетерозиготном состоянии, ко всем скрещиваниям.
Определение эффекта дозы гена и использование хромосомных аберраций. Этими методами обнаруживают корреляции между уровнем экспрессии исследуемого гена и количеством конкретных хромосом в анеуплоидных линиях клеток или структурными перестройками хромосом (хромосомными мутациями – аберрациями). Анеуплоидией называют наличие у клетки, ткани или целого организма числа хромосом, не равного типичному для данного биологического вида. Хромосомные аберрации в виде транслокаций участков хромосом в гетерохроматиновые области тех же самых или других хромосом часто сопровождаются подавлением транскрипции генов, расположенных в транслоцированных участках или в хромосоме-акцепторе (мозаичный эффект положения). Изменение уровня активности исследуемого гена при конкретных транслокациях может быть основанием для картирования его в соответствующей области хромосомы.
Использование синтении. Синтения – это структурное сходство групп сцепления генов у организмов разных биологических видов. В частности, в геномах человека и мыши известно несколько десятков синтеничных групп генов. Наличие феномена синтении позволяет суживать круг поиска места локализации исследуемого гена на хромосомах, ограничивая его областью известных генов, принадлежащих к конкретной синтеничной группе.
Сегрегация генов, индуцируемая ионизирующим излучением. С помощью этого метода определяют расстояние между исследуемыми генами путем оценки вероятности их разделения (сегрегации) после облучения клеток определенной стандартной дозой ионизирующего излучения. Облученные клетки спасают от гибели гибридизацией с соматическими клетками грызунов, и у соматических гибридов в культуре определяют наличие исследуемых маркеров облученных клеток. В итоге удается сделать вывод о наличии или отсутствии сцепления (физическом расстоянии) между этими генами.
Среди других методов следует упомянуть способы, основанные на использовании для картирования генов больших фрагментов ДНК, образуемых под действием крупнощепящих рестриктаз. После расщепления геномной ДНК образующиеся фрагменты разделяют электрофорезом в импульсном электрическом поле и далее их гибридизуют по Саузерну с зондами, соответствующими картируемым генам. Если после проведения гибридизации сигналы обоих зондов локализуются на одном и том же крупном фрагменте ДНК, это говорит о тесном сцеплении таких генов.