
Чем обусловлен различный характер сцепления генов
Хромосомная теория
Хромосомная теория наследственности
Концепция данной теории заключается в том, что передача наследственной информации в ряду поколений осуществляется путем передачи хромосом, в которых в определенной линейной последовательности расположены гены.
Данная теория была сформулирована в начале XX века. Значительный вклад в ее развитие внес американский генетик Томас Морган.
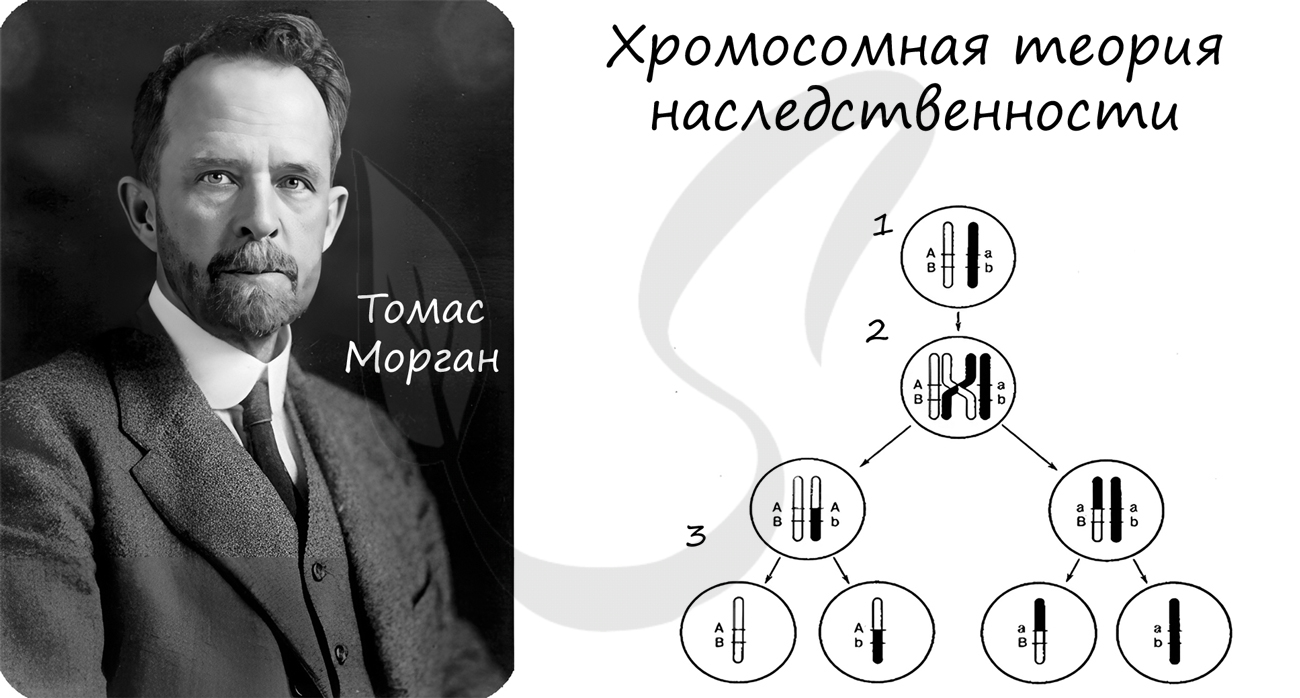
Рекомендую осознать и запомнить следующие положения хромосомной теории:
- Гены расположены в хромосомах в линейном порядке
- Каждый ген занимает в хромосоме определенное место — локус
- Гены, расположенные в одной хромосоме, образуют группу сцепления
- Сцепление генов может нарушаться в результате кроссинговера
- Частота кроссинговера между генами прямо пропорциональна расстоянию между ними
- Расстояние между генами измеряется в морганидах (1 морганида — 1% кроссинговера)
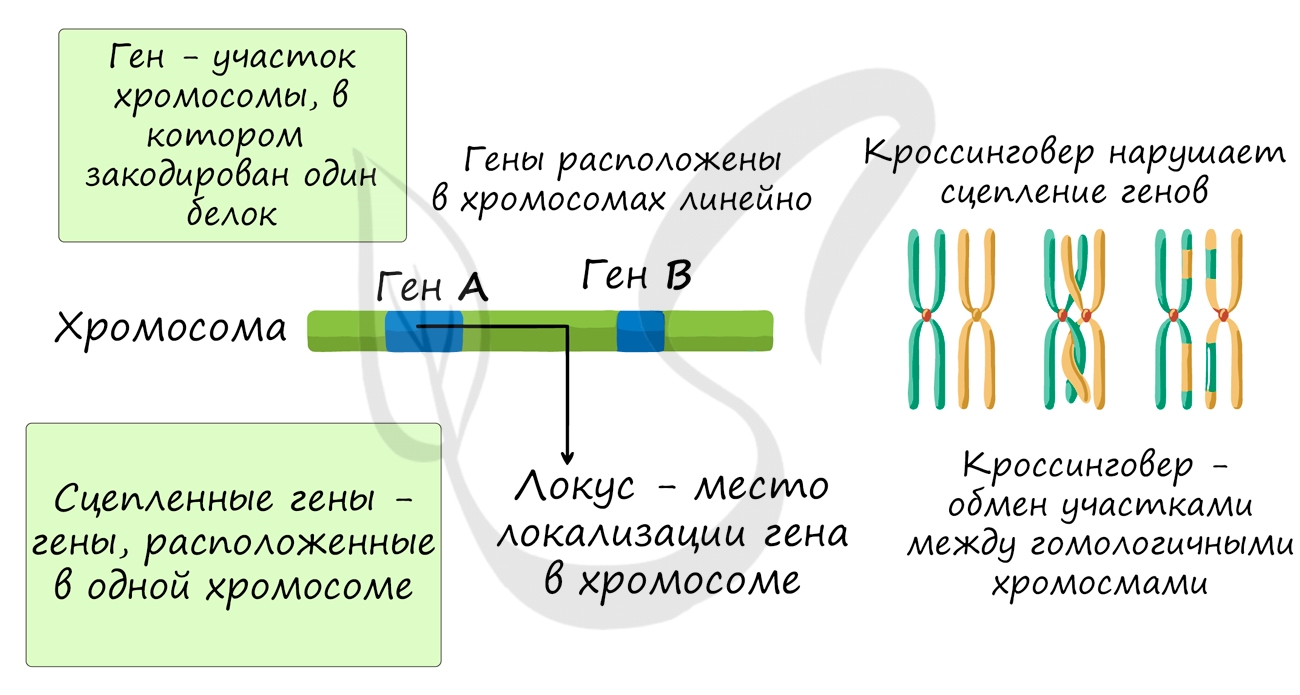
Группы сцепления
В предыдущей статье были раскрыты суть и применение в задачах III закона Менделя, закона независимого наследования, в основе которого лежат гены, расположенные в разных хромосомах. Но что если гены лежат в одной хромосоме? Такие гены образуют группу сцепления, в этом случае говорят о сцепленном наследовании.
Группа сцепления — совокупность всех генов, расположенных в одной хромосоме, вследствие чего они наследуются совместно. Число групп сцепления равно гаплоидному набору хромосом: у женщины 23 группы сцепления (23 пара — половые хромосомы XX), а у мужчины — 24 группы сцепления (X и Y представляют собой две отдельные группы).
Сцепление генов
Томас Морган в своих экспериментах изучал наследование признаков плодовых мушек дрозофил: серый (A) — черный (a) цвет тела, длинные (B) — зачаточные (b) крылья. В первом эксперименте Морган скрестил чистые линии плодовых мушек: серых с длинными крыльями (AABB) и черных с зачаточными (aabb).

Только что вы видели первый закон Менделя (единообразия) в действии, правда, в несколько ином варианте — при дигибридном скрещивании. Но суть та же: в первом поколении все особи получаются единообразны по исследуемому признаку, с генотипом AaBb — с серым телом и длинными крыльями.
Далее Морган применил анализирующее скрещивание. Полученную в первом поколении дигетерозиготу (AaBb) он скрестил с черной особью с зачаточными крыльями (aabb). Результат весьма удивил Моргана и его коллег: помимо потомства с ожидаемыми фенотипами (серое тело + длинные крылья, черное тело + зачаточные крылья) были получены особи со смешанными признаками.

Потомство со смешанными признаками подразумевает под собой особи Aabb (серое тело + зачаточные крылья) и aaBb (черные тело + длинные крылья). Но откуда они могли взяться, если гены A и B находятся в одной хромосоме? Значит, образовались еще какие-то дополнительные гаметы, помимо AB и ab?
Объясняя полученные в потомстве фенотипы, которые содержали смешанные признаки, Томас Морган пришел к выводу, что между гомологичными хромосомами произошел кроссинговер, в результате которого образовались гаметы Ab, aB — кроссоверные гаметы.
Очевидно, что в данном случае расстояние между генами A и B было 17 морганид, так как каждой кроссоверной гаметы (соответственно и особей) образовалось по 8.5%. Не забывайте, что процент кроссинговера равен расстоянию между генами. Поскольку расстояние было 17 морганид = 17%, то на каждую из кроссоверных гамет приходится половина — 8.5%
Пример решения генетической задачи №1
«Катаракта и полидактилия у человека обусловлены доминантными аутосомными генами, расположенными в одной хромосоме. Гены полностью сцеплены. Какова вероятность родить здорового ребенка в семье, где муж нормален, жена гетерозиготна по обоим признакам, мать жены также страдала обеими аномалиями, а отец был нормален».
Очень важно обратить внимание на то, что «гены полностью сцеплены» — это говорит об отсутствии кроссинговера, и то, что мы заметили это, обеспечивает верное решение задачи.

Самое главное, что вам следует усвоить: поскольку гены полностью сцеплены (кроссинговер отсутствует), женщина с генотипом AaBb может образовать только два типа гамет — AB, ab. Кроссоверные гаметы (Ab, aB) не образуются. Всего возможных генотипов потомков получается два, из которых здоров только один — aabb. Шанс родить здорового ребенка в такой семье ½ (50%).
Пример решения генетической задачи №2
«Гены доминантных признаков катаракты и эллиптоцитоза локализованы в 1-й аутосоме. Гены неполностью сцеплены. Женщина, болеющая катарактой и эллиптоцитозом, отец которой был здоров, выходит замуж за здорового мужчину. Определите возможные фенотипы потомства и вероятность рождения больного обеими аномалиями ребенка в этой семье».
Ключевые слова в тексте этой задачи, на которые следует обратить внимание: «гены неполностью сцеплены». Это означает, что между ними происходит кроссинговер.
Генотип женщины остается неясен из текста задачи. Раз она больна, то он может быть: AaBb, AABB, AABb, AaBB. Однако в тексте дано то, что развеет сомнения: «отец которой был здоров». Если ее отец был здоров, то его генотип был aabb, значит он передал дочери гамету ab. Теперь становится очевидно, что генотип дочери AaBb — она дигетерозиготна.

В данном случае между генами A и B произошел кроссинговер, их сцепление нарушилось. В результате образовались кроссоверные гаметы Ab, aB — которые привели к образованию особей с со смешанными признаками (Aabb, aaBb). Вероятность рождения в этой семье ребенка, больного обеими аномалиями, составляет ¼ (25%).
Наследование, сцепленное с полом
Половые хромосомы X и Y определяют пол человека. Генотип XX характерен для женщин, а XY — для мужчин. Мужская Y-хромосома не содержит аллелей многих генов, которые есть в X-хромосоме, вследствие этого наследственными заболеваниями, сцепленными с полом, чаще болеют мужчины.
Природа, несомненно, бережет женских особей. Женщины имеют две гомологичные хромосомы XX, и если ген наследственного заболевания попал в одну из X-хромосом, то чаще всего в другой X-хромосоме окажется «здоровый» ген, доминантный, которой подавит действие рецессивного гена. С генетической точки зрения, женщина будет носительницей заболевания, может его передать по поколению, но сама болеть не будет.
У мужчин если ген заболевания оказался в X-хромосоме, то не проявиться он не может. Именно по этой причине мужчины чаще страдают дальтонизмом, гемофилией и т.д.
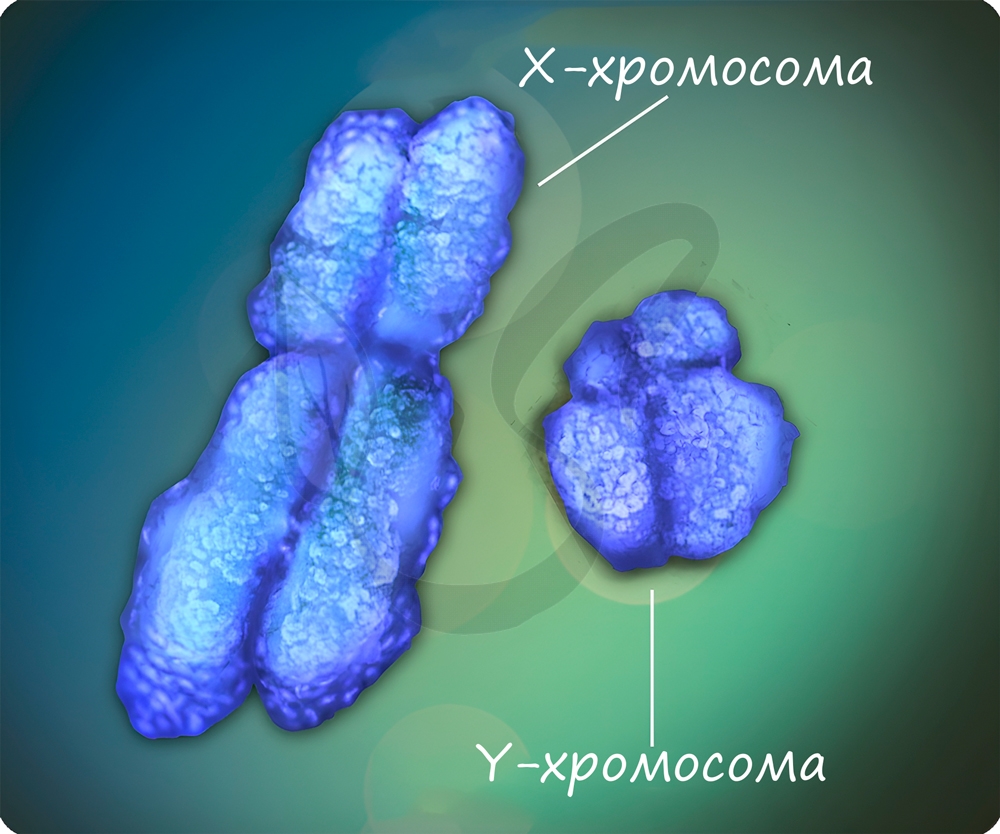
Не у всех организмов особь мужского пола характеризуется набором хромосом XY, а женского — XX. У пресмыкающихся, птиц, бабочек женские особи имеют гетерогаметный пол- XY, а мужские — XX. То же самое относится к домашним курам: петух — XX, курица — XY.
Решим несколько задач по теме наследования, сцепленного с полом. Речь в них будет идти о сцепленных с полом признаками — признаками, гены которых лежат не в аутосомах, а в гетеросомах (половых хромосомах).
Пример решения генетической задачи №3
«Рецессивный ген дальтонизма располагается в X-хромосоме. Женщина с нормальным зрением (отец был дальтоник) выходит замуж за мужчину с нормальным зрением, отец которого был дальтоником. Определите возможные фенотипы потомства».
Подробности о родословной важны и помогают заполнить белые пятна. Если отец женщины был дальтоником (X d Y), то очевидно, что он передал ей хромосому X d , так как от отца дочери всегда передается X-хромосома. Значит женщина гетерозиготна по данному признаку, а у мужчины возможен лишь один вариант здорового генотипа — X D Y. То, что его отец был дальтоником несущественно, ведь отец всегда передает сыну Y-хромосому.

Возможные фенотипы потомства:
- X D X D , X D X d — фенотипически здоровые девочки
- X D Y — здоровый мальчик
- X d Y — мальчик, который болен дальтонизмом
Пример решения генетической задачи №4
«Гипоплазия зубной эмали наследуется как сцепленный с X-хромосомой доминантный признак, шестипалость — как аутосомно-доминантный. В семье, где мать шестипалая, а у отца гипоплазия, родился пятипалый здоровый мальчик. Напишите генотипы всех членов семьи по данным признакам. Возможно ли у них рождение ребенка с двумя аномалиями одновременно?»
Ответ на вопрос: «Каковы генотипы матери и отца?» — лежат в потомстве. Пятипалый здоровый мальчик имеет генотип aaX b Y. Чтобы сформировался такой генотип, от матери должна прийти гамета aX b , а от отца — aY. Выходит, что единственно возможный генотип матери — AaX b X b , а генотип отца — aaX B Y.

Рождение ребенка с двумя аномалиями возможно — AaX B X b , вероятность такого события ¼ (25%).
Пример решения генетической задачи №5
«Рецессивные гены, кодирующие признаки дальтонизма и гемофилии, сцеплены с X-хромосомой. Мужчина с нормальным цветовым зрением и гемофилией женится на здоровой женщине, отец которой был дальтоником, но не гемофиликом. Известно, что мать женщины была гомозиготна по исследуемым признакам. Какое потомство получится от брака их дочери со здоровым мужчиной?»
Генотип мужчины вопросов не вызывает, так как единственный возможный вариант — X hD Y. Генотип женщины дает возможность узнать ее отец (X Hd Y), который передал ей гамету X Hd (отец всегда передает дочке X хромосому, а сыну — Y), следовательно, ее генотип — X HD X Hd
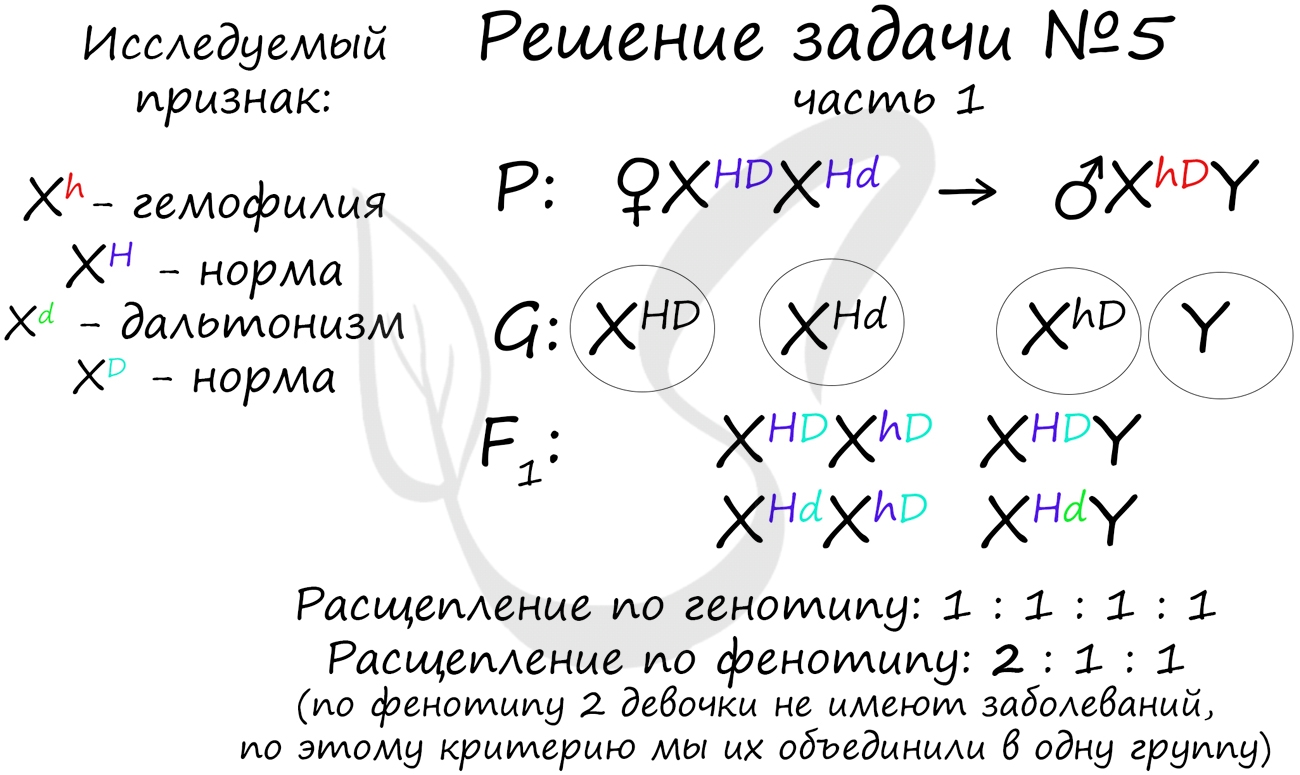
Как оказалось, возможны два варианта генотипа дочери: X HD X hD , X Hd X hD . Генотип здорового мужчины X HD Y. Следуя логике задачи, мы рассмотрим два возможных варианта брака.
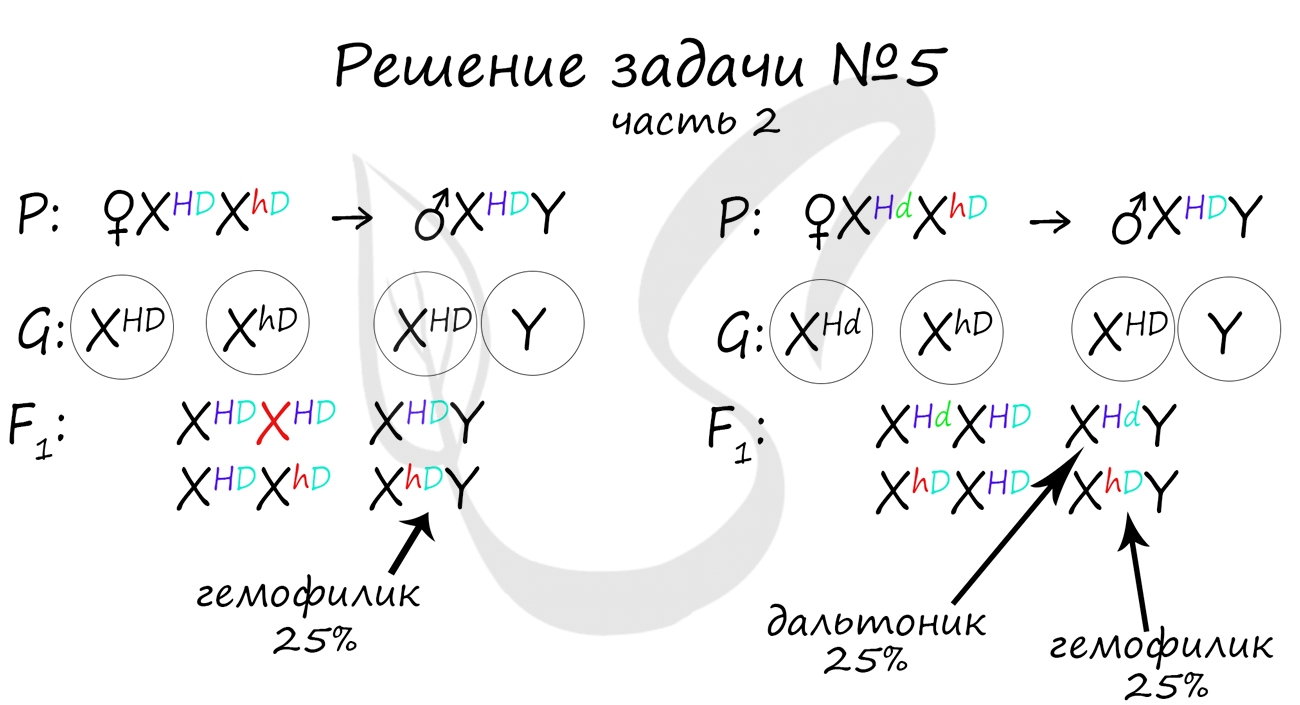
Не забывайте, что на экзамене схема задачи не является ответом. Ответ начинается только после того, как вы напишите слово «Ответ: . «. В ответе должны быть указаны все фенотипы потомства, их описание, что возможно покажется рутинными при большом числе потомков, но весьма приятным, если вы верно решили задачу и получили за нее заслуженные баллы 
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Биология. 10 класс
Хромосомная теория наследственности
Сцепленное наследование генов
Необходимо запомнить
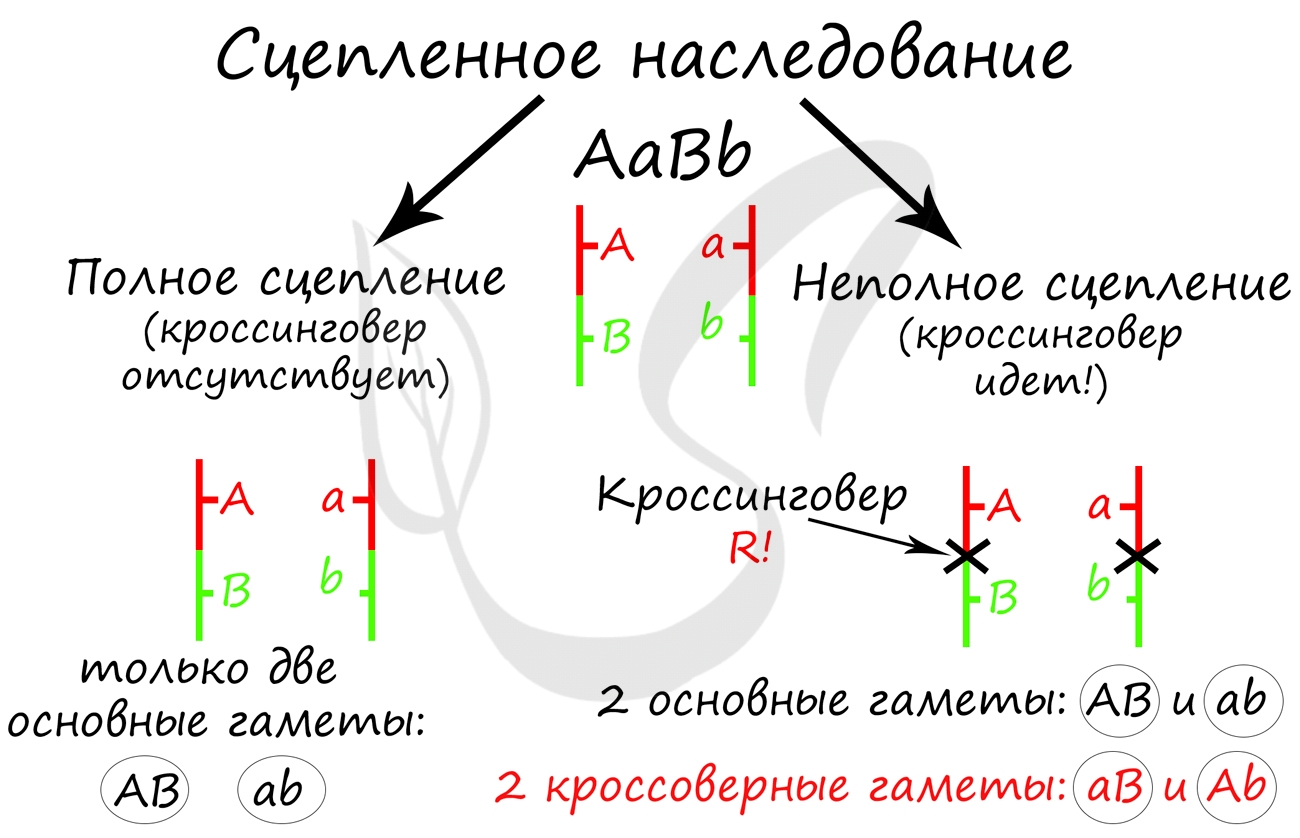
Сцепление генов – это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках её тела находится только 4 пары хромосом и имеет место высокая скорость размножения (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана). Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение.

На рисунке 1 слева : расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется.
На рисунке 1 справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В , поэтому сцепление неполное, хромосомы в гаметах образуются четырёх типов – 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах – единицах расстояния между генами, находящимися в одной хромосоме. 1 морганида соответствует 1 % кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер).
Пример, основанный на опытах Моргана

Гены, расположенные в одной хромосоме, наследуются совместно. Фенотипы А – серое тело, нормальные крылья (повторяет материнскую форму). Б – тёмное тело, короткие крылья (повторяет отцовскую форму). В – серое тело, короткие крылья (отличается от родителей). Г – тёмное тело, нормальные крылья (отличается от родителей). В и Г получены в результате кроссинговера в мейозе.
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, — доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F 1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F 2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья– сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т. е. они находятся в одной хромосоме. наследование сцепленных генов называют сцепленным наследованием.
Сцепление может нарушаться. Это доказывает наличие особей В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.

На рисунке 3 опыт Моргана отображен подробно. Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре типа гамет:

Сцепленное наследование : два гена находятся в одной хромосоме.
а) При полном сцеплении гетерозигота дает только два типа гамет.
б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью.

На вышесказанном строится хромосомная теория наследственности Моргана:
1. Гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга.
2. Гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно (т. е. в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм).
3. Новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами.
4. Учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом. За единицу расстояния между генами принята частота кроссинговера равная 1 % (морганида, сантиморган, сМ).
Задачи на нахождение расстояния между генами
Решение задачи на определение вероятности рождения здоровый и больных детей
Наследование признаков, сцепленных с полом
Сцепленными с полом называются признаки, гены которых расположены не в аутосоме (неполовой хромосоме), а в гетеросоме (половой хромосоме). Схема решения задач на наследование признаков, сцепленных с полом, иная, чем на аутосомное моногибридное скрещивание. В случае, если ген сцеплен с Х—хромосомой, он может передаваться от отца только дочерям, а от матери в равной степени и дочерям, и сыновьям. Если ген сцеплен с Х—хромосомой и является рецессивным, то у самки он проявляется только в гомозиготном состоянии. У самцов второй Х-хромосомы нет, поэтому такой ген проявляется всегда.
При решении задач этого типа используются не символы генов (А, а, В, b), как при аутосомном наследовании, а символы половых хромосом X, Y с указанием локализованных в них генов (X А , X а ).
Аномалии, сцепленные с полом, чаще контролируются рецессивными генами, локализованы в Х-хромосоме и проявляются при генотипе ХY (т. е. у самцов млекопитающих и самок птиц).
Выше были рассмотрены примеры, где ген, сцепленный с полом, располагался в Х-хромосоме, но есть гены, локализованные в Y-хромосоме. У видов, у которых мужской пол гетерогаметен, этот ген может передаваться только самцам. У человека ген одного из видов синдактилии, выражающейся в образовании перепонки между 2 и 3 пальцами на ноге, локализован на Y-хромосоме, поэтому синдактилия возникает только у мужчин. Известна еще одна аномалия – гипертрихоз края ушной раковины (ряды волос на ухе), передающиеся по такому же механизму. В изучаемой семье с этой аномалией она передавалась в пяти поколениях по мужской линии.
Биология. 11 класс
§ 34. Хромосомная теория наследственности
Как уже отмечалось, Г. Мендель, установивший важнейшие закономерности наследования, выдвинул предположение о существовании особых наследственных факторов, контролирующих признаки живых организмов. Однако материальная природа этих факторов, впоследствии названных генами, долгое время оставалась невыясненной. Лишь в начале ХХ в., после переоткрытия законов Г. Менделя, цитологи обратили внимание на связь гипотетических наследственных факторов с поведением хромосом при мейозе и оплодотворении. В 1902—1903 гг. немецкий эмбриолог Т. Бовери и американский цитолог У. Саттон (фамилии приведены не для запоминания) независимо друг от друга пришли к выводу, что именно хромосомы и являются носителями менделевских факторов. Однако ученые не смогли предоставить четких доказательств своей правоты, поэтому их предположение в течение нескольких лет так и оставалось «хромосомной гипотезой».
Хромосомная теория наследственности. Сцепленное наследование . Убедительные доказательства того, что гены располагаются в хромосомах, были получены в 1910 г. американским генетиком, будущим лауреатом Нобелевской премии (1933 г.) Т. Морганом. Многочисленные эксперименты Моргана и его сотрудников привели к ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из положений этой теории можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив только в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом — 23 пары (2n = 46), у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2n = 8). Следовательно, каждая хромосома содержит множество генов. Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Такое совместное наследование генов Т. Морган назвал сцепленным наследованием (в отличие от независимого). Гомологичные хромосомы каждой пары содержат гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы — 4.
Вам известно, что при независимом наследовании дигетерозиготная особь, например , образует четыре типа гамет в равном соотношении, т. е. по 25 %: , , и . Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота будет производить лишь два типа гамет: 50 % и 50 % . Обратите внимание на то, что сцепленные гены записываются в одну хромосому.
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два типа гамет, а четыре. Кроме ожидаемых и , формируются также гаметы с новыми комбинациями генов: и , но в меньшем процентном соотношении, чем при независимом наследовании. Рассмотрим один из опытов, в котором Т. Морган изучал наследование сцепленных генов у дрозофилы.
 *Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.*
*Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.*
Чистую линию дрозофил, имеющих серое тело и нормальные (длинные) крылья, скрестили с чистой линией, особи которой имели черное тело и зачаточные крылья (рис. 34.2). Полученные гибриды первого поколения в соответствии с первым законом Менделя были единообразными — серыми с нормально развитыми крыльями. Следовательно, у дрозофилы серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.
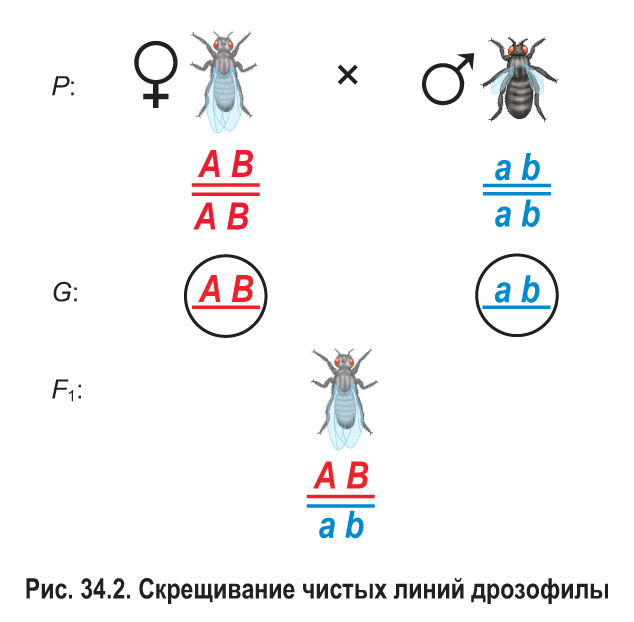
Затем было проведено анализирующее скрещивание (рис. 34.3). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.
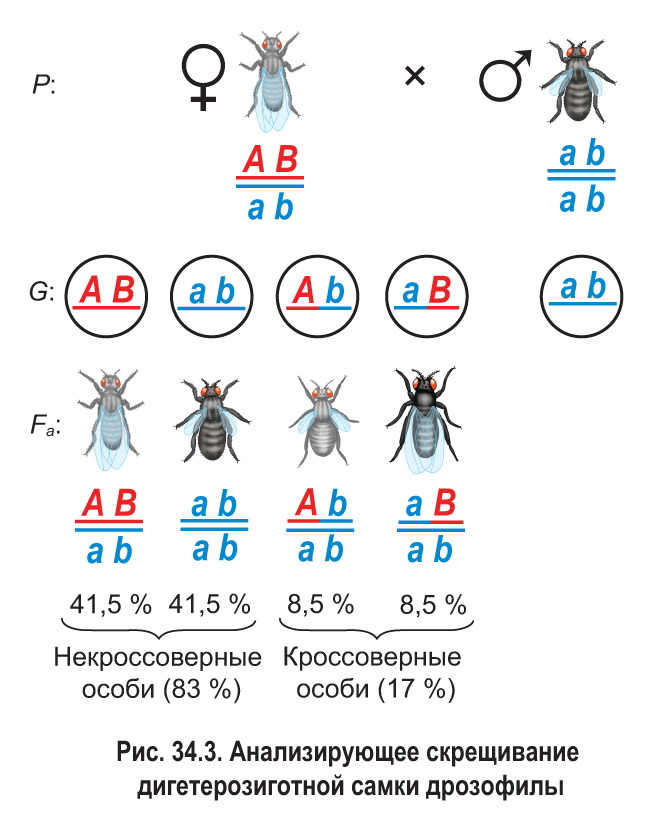
Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов, самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше ( и вместе составили 83 %), чем с новыми их комбинациями (сумма и равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы знаете, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы , которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными. В рассмотренном эксперименте гаметы и являлись кроссоверными, а гаметы и — некроссоверными (см. рис. 34.3).
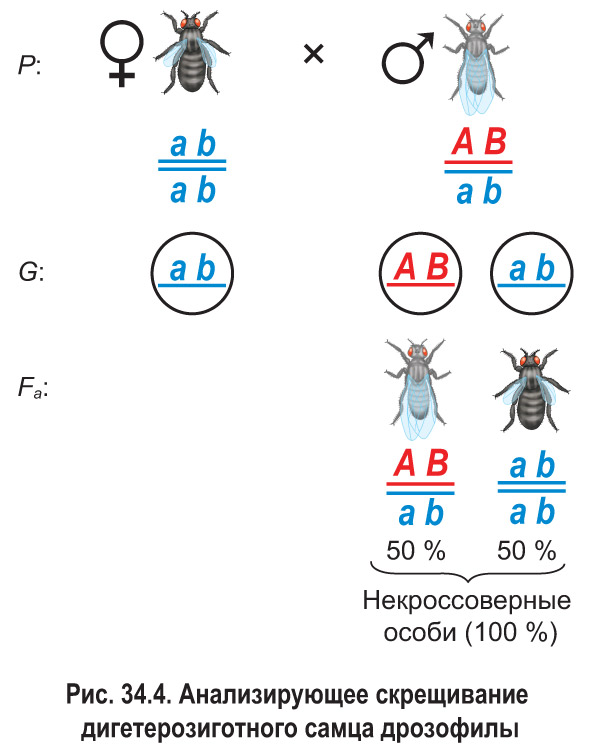
*Т. Морган подвергал анализирующему скрещиванию не только дигетерозиготных самок дрозофилы, но и самцов, т. е. проводил реципрóкные скрещивания. Так называют пару скрещиваний, в одном из которых организмы с исследуемыми признаками (или признаком) используются в качестве материнских, а в другом — в качестве отцовских. Оказалось, что реципрокные анализирующие скрещивания дигетерозиготных особей дрозофилы дают разные результаты.
В потомстве дигетерозиготного самца наблюдалось лишь два фенотипических класса (рис. 34.4). Половину потомков составляли особи с серым телом и нормальными крыльями, вторую половину — особи с черным телом и зачаточными крыльями. Это свидетельствовало об отсутствии кроссинговера между сцепленными генами или, иначе говоря, о полном (абсолютном) сцеплении генов. Выяснилось, что у самцов дрозофилы в ходе мейоза действительно не происходит кроссинговер между гомологичными хромосомами. Впоследствии подобное явление было обнаружено у самцов некоторых других видов двукрылых, а также у самок тутового шелкопряда. Однако полное сцепление генов наблюдается в природе очень редко, как исключение из общего правила.*
Кроссинговер между сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB = 17 %.
Дальнейшие исследования, проведенные Т. Морганом и сотрудниками его лаборатории, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними. Чем объясняется эта закономерность?
В профазе I мейоза при конъюгации гомологичных хромосом взаимный обмен участками между хроматидами осуществляется произвольно, в любой их точке. Рассмотрим рисунок 34.5.
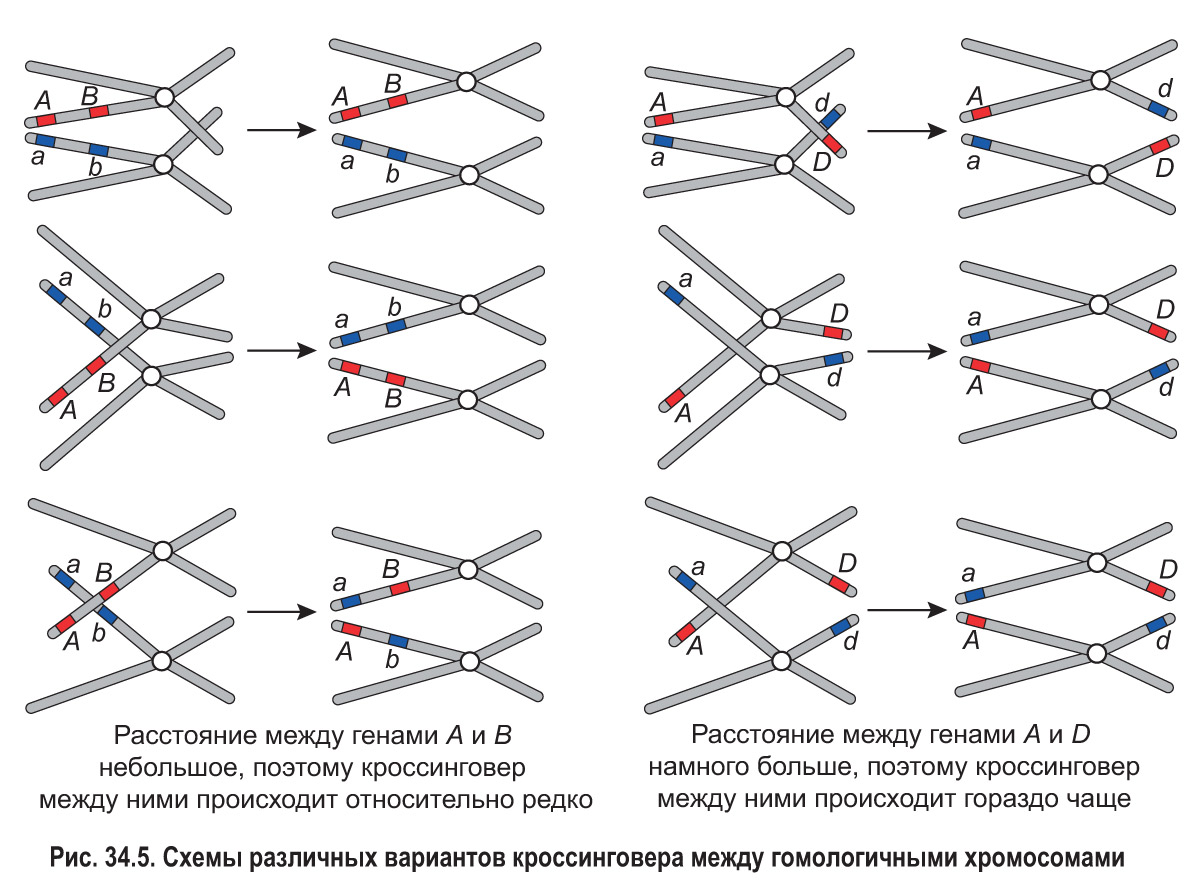
Гены А и В (или а и b) находятся сравнительно близко друг к другу. Вероятность того, что обмен произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды совершат обмен в какой-либо точке между этими генами, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название морганида или, что то же самое, сантиморган (сМ).
Морганида (сантиморган, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Кроссинговер играет очень важную биологическую роль. В результате этого процесса у потомков возникают новые комбинации родительских генов, что повышает генетическое разнообразие организмов и расширяет возможности их адаптации к различным условиям окружающей среды.
Сцепленное наследование. Группы сцепления. Хромосомная теория наследственности.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Вместе с признаками, которые наследуются независимо, должны существовать и такие, которые наследуются сцеплено друг с другом, так как они определяются генами, расположенными в одной хромосоме. Такие гены образуют группу сцепления. Количество групп сцепления в организмах определенного вида равно количеству хромосом в гаплоидном наборе (например, у дрозофилы 1пара = 4, у человека 1пара = 23).
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
(Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
Кроссинговер — обмен участками гомологичных хромосом в процессе клеточного деления, преимущественно в профазе первого мейотического разделения, иногда в митозе. Опытами Т. Моргана, К. Бриджеса и А. Стертеванта было показано, что нет абсолютно полного сцепления генов, при котором гены передавались бы всегда вместе. Вероятность того, что два гена, локализованные в одной хромосоме, не разойдутся в процессе мейоза, колеблется в пределах 1-0,5. В природе преобладает неполное сцепление, обусловленное перекрёстком гомологичных хромосом и рекомбинацией генов. Цитологическая картина кроссинговера была впервые описана датским ученым Ф. Янсенсом .
Кроссинговер проявляется только тогда, когда гены находятся в гетерозиготном состоянии (АВ / ав). Если гены находятся в гомозиготном состоянии (АВ / АВ или аВ/аВ), обмен идентичными участками не дает новых комбинаций генов в гаметах и в поколении. Частота (процент) перекрёстка между генами зависит от расстояния между ними: чем дальше они располагаются друг от друга, тем чаще происходит кроссинговер. Т. Морган предложил расстояние между генами измерять кроссинговером в процентах, по формуле:
N1/N2 X 100 = % кроссинговера,
где N1 — общее число особей в F;
N2 — суммарное число кроссоверных особей.
Отрезок хромосомы, на котором осуществляется 1% кроссинговера, равна одной морганиде (условная мера расстояния между генами). Частоту кроссинговера используют для того, чтобы определить взаимное расположение генов и расстояние между ними. Для построения генетической карты человека пользуются новыми технологиями, кроме того построены цитогенетические карты хромосом.
Различают несколько типов кроссинговера: двойной, множественный (сложный), неправильный, неровный.
Кроссинговер приводит к новому сочетанию генов, вызывает изменение фенотипа. Кроме того, он наряду с мутациями является важным фактором эволюции организмов.)
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
· гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
· каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
· гены расположены в хромосомах в определенной линейной последовательности;
· гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
· сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
· каждый вид имеет характерный только для него набор хромосом — кариотип.
Наследование пола и признаков, сцепленных с полом. Половые хромосомы и их роль в детерминации пола.Наследование пола. Пол особи — это сложный признак, формируемый как действием генов, так и условиями развития. У человека одна из 23 пар хромосом — половые хромосомы, обозначаемые как X и Y. Женщины — гомогаметный пол, т.е. имеют две X-хромосомы, одну — полученную от матери, а другую — от отца. Мужчины — гетерогаметный пол, имеют одну X- одну Y-хромосому, причем X передается от матери, а Y — от отца. Заметим, что гетерогаметный пол не всегда обязательно мужской; например, у птиц это самки, в то время как самцы гомогаметны. Имеются и другие механизмы детерминации пола. Так, у ряда насекомых Y-хромосома отсутствует. При этом один из полов развивается при наличии двух X-хромосом, а другой — при наличии одной X-хромосомы. У некоторых насекомых пол определяется соотношением числа аутосом и половых хромосом. У ряда животных может происходить т.н. переопределение пола, когда в зависимости от факторов внешней среды зигота развивается либо в самку, либо в самца. Развитие пола у растений имеет столь же разнообразные генетические механизмы, как и у животных.
Признаки, сцепленные с X-хромосомой. Если ген находится в половой хромосоме (его называют сцепленным с полом), то проявление его у потомков следует иным, чем для аутосомых генов, правилам. Рассмотрим гены, находящиеся в X-хромосоме. Дочь наследует две X-хромосомы: одну — от матери, а другую — от отца. Сын же имеет только одну X-хромосому — от матери; от отца же он получает Y-хромосому. Поэтому отец передает гены, имеющиеся в его X-хромосоме, только своей дочери, сын же их получить не может. Поскольку X-хромосома более «богата» генами по сравнению с Y-хромосомой, то в этом смысле дочь генетически более схожа с отцом, чем сын; сын же более схож с матерью, чем с отцом.
Один из исторически наиболее известных сцепленных с полом признаков у человека — это гемофилия, приводящая к тяжелым кровотечениям при малейших порезах и обширным гематомам при ушибах. Она вызывается рецессивным дефектным аллелем 0, блокирующим синтез белка, необходимого для свертывания крови. Ген этого белка локализован в Х-хромосоме. Гетерозиготная женщина +0 (+ означает нормальный активный аллель, доминантный по отношению к аллелю гемофилии 0) не заболевает гемофилией, и ее дочери тоже, если у отца нет этой патологии. Однако ее сын может получить аллель 0, и тогда у него развивается гемофилия. Рецессивные заболевания, вызываемые генами X-хромосомы, намного реже поражают женщин, чем мужчин, поскольку у них заболевание проявляется только при гомозиготности — наличии рецессивного аллеля в каждой из двух гомологичных X-хромосом; мужчины заболевают во всех случаях, когда их единственная X-хромосома несет дефектный аллель.
Сцепление с Y-хромосомой. Сведения о генах, находящихся в Y-хромосоме, весьма скудны. Предполагается, что она практически не несет генов, обусловливающих синтез белков, необходимых для функционирования клетки. Но она играет ключевую роль в развитии мужского фенотипа. Отсутствие Y-хромосомы при наличии только одной X-хромосомы приводит к т.н. синдрому Тернера: развитию женского фенотипа с плохо развитыми первичными и вторичными половыми признаками и другими отклонениями от нормы. Встречаются мужчины с добавочной Y-хромосомой (XYY); они высокого роста, агрессивны и нередко аномального поведения. В Y-хромосоме выявлено несколько генов, ответственных за регуляцию синтеза специфических ферментов и гормонов, и нарушения в них приводят к патологиям полового развития. Имеется ряд морфологических признаков, которые, как полагают, определяются генами Y-хромосомы; среди них — развитие волосяного покрова ушей. Подобного рода признаки передаются только по мужской линии: от отца к сыну.
Генетическая детерминация пола, определяемая набором половых хромосом, поддерживает равное воспроизводство самок и самцов. Действительно, женские яйцеклетки содержат только X-хромосому, поскольку женщины имеют генотип XX по половым хромосомам. Генотип же мужчин — XY, и потому рождение девочки или мальчика в каждом конкретном случае определяется тем, несет ли спермий X- или Y-хромосому. Поскольку же в процессе мейоза хромосомы имеют равные шансы попасть в гамету, то половина гамет, производимых индивидами мужского пола, содержит X-, а половина — Y-хромосому. Поэтому половина потомков ожидается одного пола, а половина — другого.
Следует подчеркнуть, что предсказать заранее рождение мальчика или девочки невозможно, поскольку невозможно предугадать, какая мужская половая клетка будет участвовать в оплодотворении яйцеклетки: несущая X- или Y-хромосому. Поэтому наличие большего или меньшего числа мальчиков в семье — дело случая.
Биология в лицее
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ 







Site biology teachers lyceum № 2 Voronezh city, Russian Federation
В начале XX в., когда генетики стали проводить множество экспериментов по скрещиванию на самых различных объектах (кукуруза, томаты, мыши, мушки дрозофилы, куры и др.), обнаружилось, что не всегда проявляются закономерности, установленные Менделем. Например, не во всех парах аллелей наблюдается доминирование. Вместо него возникают промежуточные генотипы, в которых участвуют обе аллели. Обнаруживается также много пар генов, не подчиняющихся закону независимого наследования генов, особенно если пара аллельных генов находится в одной и той же хромосоме, т. е. гены как бы сцеплены друг с другом. Такие гены стали называть сцепленными.
Механизм наследования сцепленных генов, а также местоположение некоторых сцепленных генов установил американский генетик и эмбриолог Т. Морган. Он показал, что закон независимого наследования, сформулированный Менделем, действителен только в тех случаях, когда гены, несущие независимые признаки, локализованы в разных негомологичных хромосомах. Если же гены находятся в одной и той же хромосоме, то наследование признаков происходит совместно, т. е. сцепленно. Это явление стали называть сцепленным наследованием, а также законом сцепления или законом Моргана.
Закон сцепления гласит: сцепленные гены, расположеные в одной хромосоме, наследуются совместно (сцепленно) .
Примеров сцепленного наследования генов известно очень много. Например, у кукурузы окраска семян и характер их поверхности (гладкие или морщинистые), сцепленные между собой, наследуются совместно. У душистого горошка (Lathyrus odoratus) сцепленно наследуются окраска цветков и форма пыльцы.
Все гены одной хромосомы образуют единый комплекс – группу сцепления. Они обычно попадают в одну половую клетку – гамету и наследуются вместе.
Группа сцепления — все гены одной хромосомы. Число групп сцепления равно количеству хромосом в гаплоидном наборе. Например, у человека 46 хромосом — 23 группы сцепления, у гороха 14 хромосом — 7 групп сцепления, у плодовой мушки дрозофилы 8 хромосом — 4 группы сцепления.
Гены, входящие в группу сцепления, не подчиняются третьему закону Менделя о независимом наследовании. Однако полное сцепление генов встречается редко. Если гены располагаются близко друг к другу, то вероятность перекреста хромосом мала и они могут долго оставаться в одной хромосоме, а потому будут передаваться по наследству вместе. Если же расстояние между двумя генами на хромосоме велико, то существует большая доля вероятности, что они могут разойтись по разным гомологичным хромосомам. В этом случае гены подчиняются закону независимого наследования.
Таким образом, третий закон Менделя отражает частое, но не абсолютное явление в наследовании признаков.
Основные доказательства передачи наслед-ственности были получены в экспериментах Моргана и его сотрудников.
Таким образом, сцепленное наследование — явление совместного наследования генов, локализованных в одной хромосоме.
«Полное и неполное сцепление генов. Генетические карты хромосом».
Поурочное планирование 10 класс
Тема: «Полное и неполное сцепление генов. Генетические карты хромосом».
Цель урока: ознакомить учащихся с процессом полного и неполного сцепления генов; и сформировать представление о генетических картах хромосом.
Учебно – воспитательные задачи:
Раскрыть сущность явления сцепленного наследования генов.
Сформировать знания об основных положениях закона Т. Моргана.
Познакомить с принципом составления генетических карт
Развивать логическое мышления учащихся.
Оборудование, наглядные пособия : таблицы по общей биологии, иллюстрирующие сцепленное наследование генов и признаков, презентация к уроку, задачи на закрепление нового материала.
Тип урока: Урок изучения нового материала.
Методы: объяснительно — иллюстративный.
I Организационный момент
Проверка личного состава учащихся и визуальной готовности класса к уроку.
II Проверка знаний учащихся
1. Назовите три закона Г. Менделя?
2. Каких правил придерживался Г. Мендель при проведении своих опытов?
3. Сформулируйте закон чистоты гамет. Кому принадлежит открытие этого закона?
4. Всегда ли признаки можно чётко разделить на доминантные и рецессивные?
5. Какое название получило это явление?
6. Всегда ли по фенотипу можно определить, какие гены содержит данная особь? Приведите пример.
7. Можно ли установить генотип особей, которые не различаются по фенотипу? Какой метод используют для этого?
8. Какими особенностями характеризуется дигибридное скрещивание?
Молодцы! С этим этапом работы Вы справились
III Изучение нового материала:
Сцепленное наследование генов
Г.Мендель проследил наследование семи пар признаков у гороха. Многие исследователи, повторяя опыты Менделя, подтвердили открытые им законы. Было признано, что эти законы носят всеобщий характер. Однако в 1906 г. английские генетики В.Бэтсон и Р.Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве. Потомки всегда повторяли признаки родительских форм. Постепенно факты исключений из третьего закона Менделя накапливались. Стало ясно, что не для всех генов характерно независимое распределение в потомстве и свободное комбинирование.
Любой организм обладает многообразием морфологических, физиологических, биохимических и прочих признаков и свойств, причем каждый признак или свойство контролируется одним или несколькими генами, локализованными в хромосомах.
Однако если число генов организма огромно и может исчисляться десятками тысяч, то число хромосом сравнительно невелико и, как правило, измеряется несколькими десятками. Поэтому в каждой паре хромосом локализованы сотни и тысячи аллельных генов, образующих группы сцепления.
Установлено полное соответствие между числом групп сцепления и числом пар хромосом. Например, у кукурузы набор хромосом 2n = 20 и 10 групп сцепления, а у дрозофилы 2n = 8 и 4 группы сцепления, то есть число групп сцепления равно гаплоидному набору хромосом.
Закон Томаса Моргана
Гены, локализованные в одной хромосоме, передаются совместно, и способ их наследования отличается от наследования генов, локализованных в разных парах гомологичных хромосом.
Так, например, при независимом распределении хромосом дигибрид АаВb образует четыре типа гамет ( АВ , аВ , Аb , аb ), а при условии полного сцепления такой же дигибрид даст только два типа гамет ( АВ и аb ), так как эти гены расположены в одной хромосоме.
Разработка проблемы сцепленного наследования генов принадлежит школе Т.Моргана (1866–1945). Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила. Мушка каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка хорошо различимы – у самца брюшко меньше и темнее. Кроме того, они имеют различия по многочисленным признакам и могут размножаться в пробирках на дешевой питательной среде.
Изучая закономерности наследования генов, локализанных в одной и той же хромосоме , Морган пришел к выводу, что они наследуются сцепленно . Это и есть закон Т.Моргана.
Кроссинговер — процесс обмена участками гомологичных хромосом во время конъюгации в профазе I мейоза. Помимо мейотического, описан также митотический кроссинговер. Хромосома разделяется на эти участки в определённых точках, одних и тех же для одного вида, что может быть определением вида на генетическом уровне, месторасположение этих точек задаётся единственным геном.
Поскольку кроссинговер вносит возмущения в картину сцепленного наследования, его удалось использовать для картирования «групп сцепления» (хромосом). Возможность картирования была основана на предположении о том, что, чем чаще наблюдается кроссинговер между двумя генами , тем дальше друг от друга расположены эти гены в группе сцепления и тем чаще будут наблюдаться отклонения от сцепленного наследования.
Полное и неполное сцепление
Для определения типа наследования двух пар генов (сцепленное или независимое) необходимо провести анализирующее скрещивание и по его результатам сделать вывод о характере наследования генов. Рассмотрим три возможных варианта результатов анализирующего скрещивания.
1) Независимое наследование .

Если в результате анализирующего скрещивания среди гибридов образуется четыре класса фенотипов, значит, гены наследуются независимо.
2) Полное сцепление генов .

При полном сцеплении генов А и В по результатам анализирующего скрещивания обнаруживают-
ся два фенотипических класса гибридов, полностью копирующих родителей.
3) Неполное сцепление генов .

В случае неполного сцепления генов А и В при анализирующем скрещивании появляются четыре фенотипа, два из которых имеют новое сочетание генов: Аb ‖ аb ; аВ ‖ аb . Появление подобных форм свидетельствует о том, что дигибрид с гаметами АВ │ и аb │ образует кроссоверные гаметы Аb │ и аВ │. Появление таких гамет возможно только в результате обмена участками гомологичных хромосом, то есть в процессе кроссинговера. Количество кроссоверных гамет значительно меньшее, чем некроссоверных.
Частота перекреста пропорциональна расстоянию между генами. Чем ближе расположены гены в хромосоме, тем теснее сцепление между ними и тем реже они разделяются при перекресте. И наоборот, чем дальше гены отстоят друг от друга, тем слабее сцепление между ними и чаще перекрест. Следовательно, о расстоянии между генами в хромосомах можно судить по частоте перекреста.
Генетические карты
Под генетическим картированием обычно понимают определение положения какого-либо гена по отношению к другим генам.
Рассмотрим порядок составления генетических карт.
1. Установление группы сцепления (то есть определение хромосомы, в которой локализован данный ген). Для этого необходимо иметь хотя бы по одному гену-маркеру в каждой группе сцепления.
2. Нахождение места локализации исследуемого гена в хромосоме. Для этого проводится скрещивание мутантной формы с нормальной и учитывается результат кроссинговера.
3. Определение расстояния между сцепленными генами, что позволяет составлять генетические карты хромосом, на которых указаны порядок расположения генов в хромосомах и относительные расстояния их друг от друга. Чем частота кроссинговера выше, тем на большем расстоянии друг от друга располагаются гены. Если установлено, что между сцепленными генами А и В частота кроссинговера 10%, а между генами В и С – 20%, то очевидно, что расстояние ВС в 2 раза больше, чем АВ . Расстояние между генами выражается в единицах, соответствующих 1% кроссинговера. Эти единицы называют морганидами.
Таким образом, на основе данных о частоте кроссинговера составляются генетические карты.
IV Закрепление знаний
Решение генетической задачи
Самку дрозофилы, гетерозиготную по рецессивным генам темной окраски тела и миниатюрных крыльев, скрестили с самцом, имевшим темное тело и миниатюрные крылья. От этого скрещивания было получено:
– 244 мухи с темным телом и миниатюрными крыльями;
– 20 мух с серой окраской тела и миниатюрными крыльями;
– 15 мух с темной окраской тела и нормальными крыльями;
– 216 мух с серой окраской тела и нормальными крыльями.
Исходя из приведенных данных определите, являются две эти пары генов сцепленными или нет. Как гены сцеплены?
А – серое тело
а – темное тело
В – нормальные крылья
b – миниатюрные крылья
Характер наследования генов А и В – ?
Результаты расщепления среди гибридов (два фенотипических класса являются господствующими и повторяют фенотипически и генотипически родительские формы, а два других класса фенотипов представлены небольшим количеством особей) свидетельствуют о неполном сцепление генов А и В.

Ответ : гены А и В наследуются сцеплено; сцепление носит неполный характер.
VI Домашнее задание
§41, стр.161-164. Ответить на вопросы стр.165.
Задача №1. Дигетерозиготное растение гороха с гладкими семенами и усиками скрестили с растением с морщинистыми семенами без усиков. Известно, что оба доминантных гена (гладкие семена и наличие усиков) локализованы в одной хромосоме, кроссинговера не происходит. Составьте схему решения задачи. Определите генотипы родителей, фенотипы и генотипы потомства, соотношение особей с разными генотипами и фенотипами. Какой закон при этом проявляется?
А — гладкие семена, а — морщинистые семена
B — наличие усиков, b — без усиков





